


Chapitre 2
La diversité biologique : un état des lieux
© Dunod – La photocopie non autorisée est un délit.
En dépit de l’attention qui est accordée à la diversité biologique depuis une dizaine d’années par les médias et les scientifiques, nos connaissances ne permettent pas d’en dresser un état exhaustif, d’autant qu’elle n’est pas distribuée de manière uniforme sur la Planète. Nous en avons néanmoins une perception globale suffisante pour permettre de jeter les bases d’une politique de conservation, conforme aux objectifs de la Convention sur la diversité biologique.
2.1LA CLASSIFICATION DU VIVANT ET SES PRINCIPES
La classification est une manière d’organiser l’information en regroupant ce qui est similaire. On tente ainsi depuis des siècles de décrire, de nommer, de classer, de compter les espèces, et il y a différentes façons de le faire. En son temps, Aristote regroupait les humains et les oiseaux parce qu’ils marchent sur deux jambes. Aujourd’hui, les classifications sont basées sur le degré de similarité génétique entre individus et regroupent les organismes en fonction de leurs parentés phylogénétiques.
18 |
2 • La diversité biologique : un état des lieux |
|
|
La taxinomie est la discipline scientifique qui consiste à nommer, décrire et classer les êtres vivants. Cette science, très formalisée, obéit aux instructions de codes internationaux de nomenclature. La systématique quant à elle a pour objectifs l’étude de la diversité des organismes et la compréhension des relations entre les organismes vivants et fossiles, c’est-à-dire leurs degrés de parenté. Ce que l’on appelle actuellement biosystématique, est une approche moderne de la systématique qui fait appel à des informations de différentes origines: morphologie, génétique, biologie, comportement, écologie, etc.
2.1.1 Les niveaux d’organisation du monde vivant
Une des caractéristiques du monde vivant est sa structuration complexe et hiérarchisée: les atomes s’organisent en cristaux (monde inanimé) ou en molécules, et ces molécules s’organisent à leur tour en cellules capables de se reproduire (monde vivant). Les cellules peuvent s’agréger et coopérer pour constituer des organismes multicellulaires. Les individus, unis ou pluricellulaires, s’organisent en populations et en communautés multispécifiques. Si l’on prend en compte le milieu dans lequel vivent les organismes, on accède alors à des ensembles de plus en plus complexes appelés écosystèmes, paysages et biosphère. Dans cette échelle hiérarchique les éléments d’un niveau d’organisation constituent les unités élémentaires qui vont entrer dans la constitution du niveau d’organisation supérieur. À chaque étape, émergent des structures et des propriétés nouvelles qui sont le résultat des interactions entre les éléments du niveau inférieur.
L’unité élémentaire du monde vivant est l’individu, porteur d’un patrimoine génétique propre. L’ensemble de ses gènes constitue son génotype. Une bactérie contient environ 1 000 gènes, certains champignons de l’ordre de 10 000. Il y en a un peu plus de 20 000 à 25 000 chez l’homme.
L’espèce est l’ensemble des individus susceptibles d’échanges génétiques fertiles et féconds (voir paragraphe 2.1.3) Une espèce est souvent répartie en populations plus ou moins isolées qui peuvent, ou non, échanger des individus et donc des informations génétiques. La population correspond à l’ensemble des individus d’une même espèce biologique habitant un même milieu. C’est à ce niveau d’organisation que s’effectue la sélection naturelle. Des populations fragmentées interactives sont des métapopulations.
2.1 La classification du vivant et ses principes |
19 |
|
|
Les ensembles d’espèces délimités le plus souvent sur des bases taxinomiques constituent les peuplements ou les communautés. La biocénose est l’ensemble des populations d’espèces animales et végétales qui vivent dans un milieu donné.
Le terme écosystème a été introduit par Tansley en 1935 pour nommer un système écologique qui combine l’ensemble des organismes vivants et leur environnement physico-chimique. La Convention sur la diversité biologique définit l’écosystème comme un «complexe dynamique formé de communautés de plantes, d’animaux et de micro-organismes, et de leur environnement non vivant qui, par leur interaction, forment une unité fonctionnelle». Cette définition légale n’est pas différente sur le fond de ce que l’on trouve dans les traités d’écologie.
La biosphère (sensu stricto) est l’ensemble des organismes vivants qui peuplent la surface de la Terre. Néanmoins on définit aussi la biosphère (sensu lato) comme la pellicule superficielle de la planète qui renferme les êtres vivants, et dans laquelle la vie est possible en permanence. Cet espace comprend ainsi la lithosphère (écorce terrestre), l’hydrosphère (ensemble des océans et des eaux continentales), et l’atmosphère (enveloppe gazeuse de la Terre).
© Dunod – La photocopie non autorisée est un délit.
2.1.2Les hiérarchies taxinomiques:
la recherche d’un ordre évolutif et fonctionnel dans la diversité des espèces
La classification consiste à reconnaître et à définir des groupes ou taxons – c’est-à-dire un ensemble d’organismes possédant en commun au moins un caractère particulier – et à les classer dans des ensembles hiérarchisés. D’abord basée à la Renaissance sur l’idée d’une classification descendante (division a priori de grandes classes en sous-classe), la taxinomie a évolué vers une classification ascendante qui consiste à regrouper des taxa apparentés en taxa d’ordre supérieur. La classification du monde vivant postule que les espèces appartenant aux mêmes taxa partagent un certain nombre de caractéristiques morphologiques, biologiques et écologiques communes qui diffèrent de celles d’autres taxa.
Les naturalistes du monde entier utilisent un même système de nomenclature générale proposé par Linné pour nommer et classer les espèces. Dans cette classification biologique, à chaque rang de la hiérarchie correspond un nom de taxon. À la base, le système binominal est constitué d’un nom de genre suivi d’un nom d’espèce. Chacune des catégories supra-spécifiques (genre, famille, ordre, division, classe, phylum, etc.)
20 |
2 • La diversité biologique : un état des lieux |
|
|
sert à rendre compte des degrés de parenté entre les taxons de rang inférieur (voir tableau 2.1).
TABLEAU 2.1 CLASSIFICATION BIOLOGIQUE HIÉRARCHIQUE DE TROIS ESPÈCES ANIMALES.
Niveau |
Espèce 1 |
Espèce 2 |
Espèce 3 |
|
|
|
|
Domaine |
Eucaryotes |
Eucaryotes |
Eucaryotes |
|
|
|
|
Règne |
Animal |
Animal |
Animal |
|
|
|
|
Embranchement |
Arthropodes |
Arthropodes |
Chordés |
|
|
|
|
Classe |
Insectes |
Crustacés |
Mammifères |
|
|
|
|
Ordre |
Diptères |
Décapodes |
Primates |
|
|
|
|
Famille |
Nématocères |
Caridés |
Hominidés |
|
|
|
|
Genre |
Aedes |
Homarus |
Homo |
|
|
|
|
Espèce |
aegypti |
americanus |
sapiens |
|
|
|
|
On a utilisé divers critères pour hiérarchiser les taxons. La hiérarchie phénétique est fondée sur la similitude des formes ou des caractères morphologiques entre les espèces. La taxonomie numérique fait l’hypothèse que les organismes qui partagent des caractéristiques communes (traits homologues) ont une histoire évolutive similaire, sans préjuger pour autant de leur généalogie. Cependant, les convergences morphologiques au cours de l’évolution ont pu conduire à des regroupements discutables. Ainsi, les Dipneustes (poissons à poumons fonctionnels tels que le Protopterus) sont plus proches morphologiquement du saumon que de la vache, mais ils ont avec la vache un ancêtre commun plus récent qu’avec le saumon. Comment faut-il donc classer les Dipneustes?
La hiérarchie phylogénétique est fondée quant à elle sur la parenté évolutive de groupes issus d’ancêtres communs. La classification cladistique (parfois appelée classification hennigienne) part du principe qu’au cours de l’évolution une espèce ancestrale donne naissance à deux espèces filles. Un groupe d’espèce est dit monophylétique lorsqu’il dérive d’un seul ancêtre commun, alors qu’un groupe polyphylétique comprend des espèces qui présentent des ressemblances, mais ne descendent pas toutes en ligne directe d’un ancêtre commun. Chez les
2.1 La classification du vivant et ses principes |
21 |
|
|
© Dunod – La photocopie non autorisée est un délit.
Vertébrés actuels, le groupe des «poissons» est un ensemble polyphylétique. Par exemple les Actinoptérygiens (comme la truite) sont plus proches des Tétrapodes que des Chondrichtyens (raies, requins).
Les méthodes de la phylogénie moléculaire reposent sur l’hypothèse que les espèces les plus apparentées présenteront des séquences de gènes qui se ressemblent plus qu’elles ne ressemblent aux séquences du même gène des autres espèces. Le degré de différence entre les séquences de gènes dans des organismes différents peut être utilisé comme une mesure de la distance évolutive entre ces organismes. La recherche de relations de parenté entre organismes se fait ici sur les gènes et non plus sur les traits morphologiques. On peut donc comparer des organismes aussi différents que les bactéries et l’homme. Mais il ne faut jamais oublier que les phylogénies ainsi établies ne sont que des reconstructions hypothétiques du passé.
Grâce au développement des outils de la biologie moléculaire, la classification phylogénétique prend actuellement le pas sur la classification phénétique. Les premières phylogénies moléculaires apparaissent dans les années 1970 quand Carl Woese entreprend de reconstruire la phylogénie des bactéries en utilisant l’ARN ribosomique: ARNr 16S chez les bactéries, puis l’ARNr 18S chez les Eucaryotes. Auparavant, on avait commencé à développer le séquençage des protéines par électrophorèse pour déterminer la séquence des acides aminés. Une technique qui a été largement utilisée mais qui est supplantée par le séquençage des acides nucléiques, ADN et ARN.
Au gré des nouvelles recherches et découvertes, la classification peut être modifiée afin de mieux refléter les différentes lignées évolutives. Des changements sont parfois fréquents, aussi bien pour les espèces actuelles que pour les espèces fossiles. La taxinomie est donc une science en perpétuelle évolution. Par exemple, les Eucaryotes constituent un groupe très hétérogène dans lequel on distinguait quatre grands ensembles: les animaux, les plantes, les champignons et un groupe mal défini, les protistes. Les phylogénies moléculaires basées sur l’ARNr 18S ont permis de mieux comprendre les relations de parenté entre ces différentes lignées. Plusieurs «super-groupes» ont été reconnus, suggérant que la pluricellularité est apparue plusieurs fois et indépendamment au cours de l’évolution. Les plantes terrestres, les champignons, les algues brunes et les animaux ont chacun une origine évolutive distincte. Les champignons paraissent évolutivement plus proches des animaux que des plantes. Nous sommes donc (toutes proportions gardées…) plus proches d’une truffe que d’une pâquerette! Les algues ne constituent pas un groupe monophylétique. Certaines se rapprochent de plantes terrestres,

22 |
2 • La diversité biologique : un état des lieux |
|
|
d’autres sont plus proches des protistes. En outre la majorité des lignées eucaryotes n’a pas de représentants pluricellulaires, ce qui sous-entend que l’évolution n’irait pas nécessairement dans le sens de la pluricellularité.
Où placer les virus?
Il existe des êtres étranges, les virus. Ils sont constitués d’un peu de matériel génétique, ADN ou ARN, enfermé dans une enveloppe de protéines. Il n’existe pas de gènes universels communs aux organismes cellulaires et aux virus. Ils ne possèdent pas de ribosomes, et donc pas d’ARNr et ne peuvent donc être inclus dans l’arbre universel du vivant. Ils exploitent les systèmes moléculaires de leurs hôtes pour exprimer leur information génétique et se multiplier.
Le biologiste André Lwoff en a donné une définition en 1957 qui sert encore de référence: ce sont des organismes de petite taille qui n’ont qu’un type d’acide nucléique (ADN ou ARN) en guise de génome, qui ne possèdent aucune des enzymes nécessaires pour produire de l’énergie, qui sont incapables de se multiplier par division, et qui sont des parasites intracellulaires obligatoires… Malgré tout cela, il existe une très grande diversité de virus qui constituent une énigme pour les scientifiques.
Après les avoir considérés comme la forme la plus élémentaire de vie, on les a même classés à une époque parmi les substances chimiques. Savoir si un virus est vivant, c’est un peu se poser la question: qu’est-ce que la vie? Ce qui est certain est que les virus échangent de l’information génétique avec les organismes vivants. Le génome d’un virus peut ajouter des gènes viraux au génome de l’hôte, et devenir en quelque sorte une partie de ce génome. Qu’ils soient ou non vivants, les virus sont donc à la frontière qui sépare le monde vivant de celui de la biochimie. Certains scientifiques les considèrent actuellement comme des parasites qui utilisent les matériaux et l’énergie d’une cellule hôte pour synthétiser les acides nucléiques et les protéines qui leur permettent de se multiplier et de se propager.
Chez les Métazoaires (figure 2.1), à la vision classique d’un arbre phylogénique à complexité croissante (figure 2.1A) est venue se substituer une vision dans laquelle tous les groupes «intermédiaires» ont disparu (figure 2.1B). Les Arthropodes seraient proches des Nématodes,
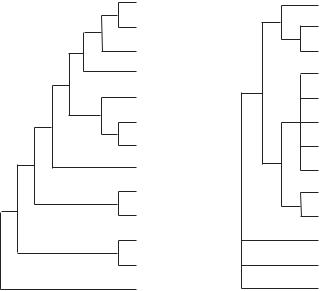
2.1 La classification du vivant et ses principes |
23 |
|
|
© Dunod – La photocopie non autorisée est un délit.
et les mollusques appartiennent à un ensemble comprenant notamment les Brachiopodes, les Annélides et les Plathelminthes. Nos insectes quant à eux sont probablement des crustacés qui se sont adaptés au milieu terrestre. La phylogénie moléculaire fait apparaître que la simplicité apparente de certains groupes considérés pendant longtemps comme primitifs (Plathelminthes, Nématodes) serait en réalité le résultat d’une simplification secondaire. De même les microsporidies, dépourvues de mitochondries, ont longtemps été considérées comme primitives, alors qu’il s’agirait en fait de lignées tardives ayant perdu secondairement leurs mitochondries.
A |
B |
Chordés |
Chordés |
Hémichordés |
Hémichordés |
Échinodermes |
Échinodermes |
Brachiopodes |
Brachiopodes |
Mollusques |
Mollusques |
Annélides |
Annélides |
Arthropodes |
Némertes |
Nématodes |
Plathelminthes |
Némertes |
Arthropodes |
Plathelminthes |
Nématodes |
Cténophores |
Cténophores |
Cnidaires |
Cnidaires |
Spongiaires |
Spongiaires |
Figure 2.1 Comparaison de deux phylogénies des métazoaires
A: la phylogénie traditionnelle où les clades successifs émergent par ordre de complexité croissante.
B: phylogénie obtenue en utilisant des séquences d’ARN 18S et de gènes Hox.
2.1.3 La notion d’espèce
La notion d’espèce est depuis longtemps un sujet de controverses, et il n’existe à l’heure actuelle aucune définition entièrement satisfaisante.
24 |
2 • La diversité biologique : un état des lieux |
|
|
Jusqu’au milieu du XVIIIe siècle, les systématiciens avaient une conception fixiste des espèces: elles étaient telles que Dieu les avait créées, en nombre limité. Le but de la taxinomie était alors d’inventorier toutes les formes de vie existantes et de décrire leurs caractères spécifiques. Linné a formalisé cette conception en matérialisant l’espèce par un individu type (holotype): l’espèce est un ensemble d’individus identiques entre eux, et avec le spécimen «type», c’est-à-dire l’exemplaire ayant servi à décrire et caractériser l’espèce sur le plan morphologique. Ce type est déposé dans un Muséum où il sert de référence, ou en quelque sorte d’étalon, pour des comparaisons ultérieures.
Cette notion fixiste n’a pas résisté à la découverte des mécanismes de l’évolution (mutation, sélection, dérive génétique) et a fait place vers le milieu du XXe siècle au concept d’espèce biologique fondé non seulement sur la ressemblance, mais également sur l’interfécondité des individus constituant une population, et dont les descendants sont euxmêmes interféconds. Ainsi, l’âne et le cheval qui peuvent se reproduire sont des espèces distinctes car leurs descendants ne sont pas féconds. C’est donc l’isolement reproductif d’un groupe d’individus qui en fait une espèce, mais encore faut-il qu’on puisse le démontrer. Il est en effet matériellement impossible de croiser la plupart des formes sauvages pour vérifier, ou non, leur interfécondité potentielle. Il en résulte une difficulté évidente pour appliquer le concept d’espèce biologique. En outre, cette définition ne s’applique en toute rigueur qu’aux espèces à reproduction bisexuée, et laisse en suspens la question pour les microorganismes. Malgré les réserves que l’on peut émettre à son sujet on continue donc, quand c’est possible, à utiliser largement la description morphologique pour identifier les espèces, en la complétant éventuellement par une description biochimique.
Au sein d’une même espèce on peut distinguer des ensembles, que l’on qualifie de sous-espèces, de races, de souches, de variétés, etc. Il n’existe pas de définition précise et universellement admise de ces catégories infraspécifiques qui peuvent être établies sur des bases morphologiques, géographiques, ou encore génétiques. Ainsi reconnaît-on parmi les nombreuses races d’animaux domestiques des formes bien différenciées sur le plan morphologique. Mais la variabilité intraspécifique peut s’exprimer de bien d’autres façons, dans le comportement reproducteur ou dans les modes de communication par exemple.
Depuis peu, les outils de la biologie moléculaire sont devenus des auxiliaires précieux pour séparer les individus appartenant à des espèces très proches sur le plan morphologique: on parle d’espèces jumelles