

PAGE PROOF: 2ND PASS
C H A P T E 19R Biomes
Our planet has a complex pattern of climates, which in turn has a major role in creating the complex pattern of vegetation and community types we find on Earth. In the previous chapter, we looked at climate and
how it affects plant physiognomy. Here we look at the resulting vegetation patterns in more detail. Ecologists divide these large-scale patterns into units called biomes: major biogeographic regions that differ from one another in the structure of their vegetation and in their dominant plant species (Clements 1916). Biomes represent the largest scale at which ecologists classify vegetation. In this chapter, we examine these broad-scale patterns, look briefly at the world’s major biomes, and briefly consider some of the ways in which human activities affect them.
Categorizing Vegetation
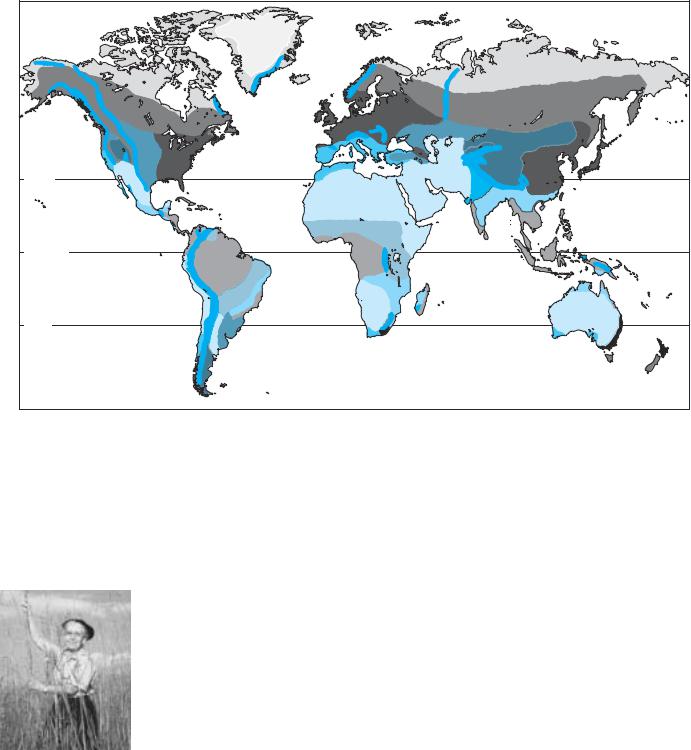
Classification of the world’s vegetation into biomes (Figure 19.1; Table 19.1) provides useful categories that describe major aspects of vegetation, such as its structure, function, and adaptations. Not only do biomes tell us a great deal about what kinds of plants might be found growing in a given area, they also indicate something about the types of animals and other organisms likely to be living there, and about the major environmental constraints on living things. Similar climates in different parts of the world contain similar biomes (Figure 19.2), although their details differ. For example, temperate evergreen forests are similar in being dominated by trees that stay green all year, but differ in being dominated by needle-leaved conifers in North America, by broad-leaved beeches in South America, and by broad-leaved eucalypts in Australia.
Biomes are defined by the physiognomy of the dominant or most obvious plants. Thus, we recognize different types of forests (e.g., evergreen or deciduous, broad-leaved or needle-leaved), shrublands, and grasslands. However, we must not lose sight of the trees for the forest: there may be a tremendous amount of variation within a given biome. In her study of communities within a single biome, the temperate deciduous forest of eastern North America, E. Lucy Braun recognized 12 different major sub-biomes and a number of minor variants (Braun 1950). Within a biome, there may be patches of vegetation that appear “not to belong,” such as riparian forests found along streams in grasslands or deserts. The occurrence of these patches reminds us that climate is only one factor in determining vegetation. Local variation in soils and topog-
380 Chapter 19
PAGE PROOF: 2ND PASS
30ºN |
Equator |
30ºS |
|
|
Tropical rainforest |
|
High mountains (taiga and alpine tundra) |
Figure 19.1 |
|
|
|
|||
|
|
Tropical deciduous forest |
|
Temperate evergreen forest |
Major biomes of the world. |
|
|
|
|
||
|
|
Thorn forest |
|
Temperate deciduous forest |
|
|
|
|
|
||
|
|
Tropical savanna |
|
Taiga |
|
|
|
|
|
||
|
|
Hot desert |
|
Arctic tundra |
|
|
|
|
|
||
|
|
Temperate shrubland |
|
Temperate grassland |
|
|
|
|
|
||
|
|
Cold desert |
|
Ice |
|
|
|
|
|
||
|
|
|
|
|
|
raphy, especially as the latter affects microclimate, can influence the type of vegetation found in an area. In turn, the animals living in a given biome are determined by the vegetation there, as well as by climate and other factors.
Biomes are useful, if somewhat arbitrary, descriptive classifications, rather than quantitative or objective categories. Bound-
aries between biomes may represent the range limits of the dominant species, whereas other species may have ranges that span those boundaries. While we draw sharp boundaries on a map, in reality the boundaries of biomes are often fuzzy. Similarly, the categories that we define ignore the fact that a particular place might not
fit easily into any given biome. Furthermore, different scientists might include a place in different biomes. As the definitions of biomes are somewhat arbitrary, they also differ in detail among scientists. Some use fewer, broader groupings, whereas others use a more detailed breakdown of vegetation types. The definitions of biomes given here may not exactly match those given in other texts. However, there is a broad consensus on the rough categories and approximate definitions of biomes.
Our use of “temperate rainforest,” for example, may not be identical to someone else’s use of that term, but it will be close. If you were to search the scientific literature, you would find many different descriptions of places called rainforests. There is, however, no single, rigorous definition of the term—it is, in fact, used to refer to forests occurring in places where rain is abundant and there is no extended dry season. Tropical rainforests occur where there is substantial year-round rain; tem-

Table 19.1 Major biomes of the world: dominant growth forms and general climatic conditionsa
|
|
Angiosperms or |
|
|
|
|
gymnosperms |
|
|
Biome |
Dominant growth form |
dominant or common |
Temperature |
Moisture |
|
|
|
|
|
Tropical rainforest |
Broad-leaved evergreen trees |
Angiosperms |
Hot |
Wet |
Tropical montane forest |
Broad-leaved evergreen trees |
Angiosperms |
Mild |
Wet |
Tropical deciduous forest |
Broad-leaved deciduous and |
Angiosperms |
Hot |
Seasonally dry |
|
semi-evergreen trees |
|
|
|
Thorn forest |
Broad-leaved deciduous trees |
Angiosperms |
Hot |
Dry |
Tropical woodland |
Broad-leaved semi-evergreen |
Angiosperms |
Hot |
Moderate |
|
trees and grasses |
|
|
|
Temperate deciduous forest |
Broad-leaved deciduous trees |
Angiosperms |
Seasonally cold |
Moderate |
Temperate rainforest |
Needle-leaved evergreen trees |
Gymnosperms |
Seasonally cool |
Wet |
Temperate evergreen forest |
Needle-leaved or broad-leaved |
Gymnosperms or |
Various |
Various |
|
evergreen trees |
angiosperms |
|
|
Temperate woodland |
Needle-leaved evergreen or |
Both |
Mild |
Moderate |
|
broad-leaved deciduous trees |
|
|
|
|
and grasses |
|
|
|
Taiga |
Needle-leaved evergreen trees |
Gymnosperms |
Cold |
Moderate |
Temperate shrubland |
Evergreen shrubs, annual forbs |
Angiosperms |
Mild |
Moderate |
Temperate grassland |
Perennial grasses |
Angiosperms |
Seasonal |
Moderate |
Tropical savanna |
Perennial grasses |
Angiosperms |
Hot |
Moderate |
Hot desert |
Shrubs, succulents, annual and |
Angiosperms |
Hot |
Dry |
|
perennial grasses, annual forbs |
|
|
|
Cold desert |
Shrubs |
Angiosperms |
Mild |
Dry |
Alpine shrubland |
Deciduous shrubs |
Angiosperms |
Cold |
Moderate |
Alpine grassland |
Perennial grasses |
Angiosperms |
Cold |
Moderate |
Tundra |
Perennial grasses, sedges, |
Angiosperms |
Cold |
Moderate |
|
shrubs, forbs |
|
|
|
aOther factors, especially seasonality, are also important.
Figure 19.2
The distribution of biomes is determined by climate, especially annual temperature and precipitation. In regions within the dashed lines, other factors—such as fire, grazing, and seasonality of precipitation—strongly affect which biome is present. Climate can also interact with factors such as soil type to determine biome distributions. (After Whittaker 1975.)
|
—15 |
|
|
|
|
|
|
|
|
|
|
|
|
—10 |
Tundra |
|
|
|
|
|
|
|
|
|
|
|
—5 |
|
Alpine |
|
Alpine shrubland |
|
|
|
|
|
|
|
|
|
|
|
Cold temperate |
|
|
|
|
|
|||
|
|
|
grassland |
|
|
|
|
|
||||
(…C) |
|
|
Cold temperate |
|
|
|
|
|
||||
0 |
|
Taiga |
|
Tropical |
|
|
|
|
|
|||
annual temperature |
|
|
|
montane forest |
|
|
|
|||||
5 |
Cold desert |
Temperate |
Temperate |
|
Te |
perate |
|
|
|
|||
woodland |
|
|
|
|
||||||||
|
|
deciduous |
|
Temperate |
|
|
|
|||||
|
|
|
everg |
en |
|
|
|
|||||
|
|
forest |
|
rainforest |
|
|
|
|||||
|
shrublandTemperate |
|
forest |
|
|
|
||||||
10 |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|||
Mean |
|
|
|
Temperate |
|
|
|
|
|
|
||
15 |
|
|
evergreen |
|
|
|
|
|
|
|||
|
Temperate |
|
|
|
|
|
|
|||||
|
|
|
|
forest |
|
|
|
Warm temperate |
||||
|
|
|
grassland |
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
||
|
20 |
|
Tropical |
|
|
|
|
|
|
|
|
|
|
|
savanna |
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
Tropical |
|
Tropical |
|
|
Tropical |
|
|
Tropical |
|
|
|
Thorn |
|
|
|
|
|
|
|||
|
25 |
|
|
deciduous |
|
rainforest |
|
|
|
|||
|
|
forest |
|
|
|
|
|
|||||
|
|
|
forest |
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|||
|
|
Hot |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|||
|
|
woodland |
|
|
|
|
|
|
|
|
||
|
30 |
desert |
|
|
|
|
|
|
|
|
|
|
|
|
50 |
100 |
150 |
200 |
250 |
|
300 |
350 |
400 |
450 |
|
|
0 |
|
|
|||||||||
Mean annual precipitation (cm)

382 Chapter 19
PAGE PROOF: 2ND PASS
Table 19.2 Primary productivity of selected major biome types
|
|
|
|
Net primary |
|
|
|
|
|
|
|
|
|
|
|
Area |
|
|
productivity per |
|
Global net primary |
Biomass per |
Total global |
||||||
|
(× 106 km2) |
|
unit area (g/m2/yr) |
|
productivity (1012 g/yr) |
unit area (kg/m) |
biomass (1012 g) |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
% of total |
|
|
|
|
|
% of total |
|
|
|
|
|
% of total |
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
terrestrial |
Normal |
|
|
|
terrestrial |
Normal |
|
|
|
terrestrial |
||
|
|
|
|
|
|
|
|
|||||||
Biome |
Amount |
area |
range |
Mean |
|
Amount |
productivity |
range |
Mean |
Amount |
biomass |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Tropical rainforest |
17.0 |
11.4 |
1000–3500 |
2200 |
37.4 |
32.5 |
|
6–80 |
45 |
|
765 |
41.7 |
||
Tropical |
7.5 |
5.0 |
1000–2500 |
1600 |
12.0 |
10.4 |
|
6–60 |
35 |
|
260 |
14.2 |
||
deciduous forest |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Temperate |
7.0 |
4.7 |
|
600–2500 |
1200 |
8.4 |
7.3 |
|
6–60 |
30 |
|
210 |
11.4 |
|
deciduous forest |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Temperate |
5.0 |
3.4 |
|
600–2500 |
1300 |
6.5 |
5.6 |
|
6–200 |
35 |
|
175 |
9.5 |
|
evergreen forest |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Woodland and |
8.5 |
5.7 |
|
250–1200 |
700 |
6.0 |
5.2 |
|
2–20 |
6 |
|
50 |
2.7 |
|
shrubland |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Taiga |
12.0 |
8.1 |
|
400–2000 |
800 |
9.6 |
8.3 |
|
6–40 |
20 |
|
240 |
13.1 |
|
Temperate |
9.0 |
6.0 |
|
200–1500 |
600 |
5.4 |
4.7 |
|
0.2–5 |
1.6 |
|
14 |
0.8 |
|
grassland |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tropical savanna |
15.0 |
10.1 |
|
200–2000 |
900 |
13.5 |
11.7 |
|
0.2–15 |
4 |
|
60 |
3.3 |
|
Desert |
18.0 |
12.1 |
|
10–250 |
90 |
1.6 |
1.4 |
|
0.1–4 |
0.7 |
|
13 |
0.7 |
|
Tundra |
8.0 |
5.4 |
|
10–400 |
140 |
1.1 |
1.0 |
|
0.1–3 |
0.6 |
|
5 |
0.3 |
|
Extreme desert, |
24.0 |
16.1 |
|
0–10 |
3 |
0.07 |
0.1 |
|
0–0.2 |
0.02 |
0.5 |
0.03 |
||
rock, sand, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
and ice |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cultivated land |
14.0 |
9.4 |
|
100–3500 |
650 |
9.1 |
7.9 |
|
0.4–12 |
1 |
|
14 |
0.8 |
|
Swamp and marsh |
2.0 |
1.3 |
|
800–1500 |
2000 |
4.0 |
3.5 |
|
3–50 |
15 |
|
30 |
1.6 |
|
Lake and stream |
2.0 |
1.3 |
|
100–1500 |
250 |
0.5 |
0.4 |
|
0–0.1 |
0.02 |
0.05 |
0.003 |
||
Total terrestrial |
149.0 |
|
|
|
773 |
115 |
|
|
|
12.3 |
|
1837 |
|
|
Total world |
510.0 |
|
|
|
333 |
170 |
|
|
|
3.6 |
|
1841 |
|
|
Source: Whittaker and Likens 1975.
perate rainforests tend to occur where there is heavy winter rain and lighter summer rain but considerable summer fog. The term “rainforest” is thus useful as a broad description, but not as a rigorously defined category or unique entity.
While we have emphasized that, scientifically, the boundaries between biomes (or communities) are arbitrary, this is not necessarily true under the law. In Australia, for example, areas designated “temperate rainforest” are legally afforded a greater measure of protection than other areas. Some eucalypt forests in very rainy places exist above an understory that is floristically a rainforest and would replace the eucalyptus canopy in about 300–500 years in the absence of fire. However, because these forests are fire-dependent, some states do not legally considered them rainforests. If the legal Australian definition were applied in North America, the redwood forests of the Pacific coast would not be classified as temperate rainforests—a categorization with which some North American scientists have agreed.
In Chapter 12, we discussed a controversy that centered on the contrasting views of Frederic Clements and Henry Gleason about the nature of communities. Clements saw communities as highly predictable entities primarily controlled by large-scale climatic patterns. In contrast, Gleason saw communities as unpredictable and variable and subject to the vagaries of dispersal, microclimatic conditions, and individualistic species distributions. One way to resolve this conflict is to recognize that each scientist emphasized a different spatial scale. Clements was focusing on patterns at the level of biomes. At that scale, we can see regular patterns and draw boundaries on our maps. Gleason was most concerned with patterns at local levels. At that scale, the boundaries blur, and we mostly see local variability. Both perspectives are valid, but each tells us different things about the world. In the following sections, we will look at vegetation patterns from a large-scale perspective as we survey the major biomes and see what distinguishes them from one another.

PAGE PROOF: 2ND PASS
Moist Tropical Forests
Tropical Rainforest
Tropical rainforests are among the most diverse and productive biomes on the planet (Figure 19.3). Although they make up only 11% of Earth’s land area, they account for over 30% of terrestrial net primary production (Table 19.2) and contain perhaps half of all living species. Tree species diversity can be as high as 300 species per 0.1 ha. Animal diversity is also high, especially that of insects and other invertebrates.
Tropical rainforests are found within a band extending from 10°N to 10°S latitude at elevations below 1 km, in areas that get more than 250 cm of rainfall each year, with some rain falling in all months. The most extensive rainforests are found in the Amazon basin of South America, the equatorial regions of western Africa, and parts of southeastern Asia. As precipitation becomes more seasonal, this biome grades into tropical semi-ever- green forests dominated by broad-leaved trees, some
(A)
Biomes 383
of which retain their leaves year-round and others that lose their leaves seasonally.
Tropical rainforests are characteristically multilayered, with two, three, or even four layers of understory trees. Tallest are the emergent trees, which are widely spaced and often more than 40 m in height, with umbrel- la-shaped canopies that extend above the general canopy of the forest. Below this is a closed canopy of trees, typically 30–40 m high. Light is readily available at the top of this layer, but greatly reduced below it. Under the closed canopy, there may be another layer of trees less than 20 m in height. Under these trees, there is often a shrub/sapling layer, and finally a sparse ground layer.
The floors of tropical rainforests can be very dark, with less than 1% of above-canopy light levels. While Hollywood movies may portray tropical rainforests as jungles with a tangle of vines and understory plants, the understory is generally sparsely vegetated with highly specialized plants capable of tolerating low light levels (see Chapter 2). The interior of a rainforest was com-
(B)
Figure 19.3
(A) The interior of a tropical rainforest in La Selva, Costa Rica. (Photograph by E. Orians.) (B) Epiphytes are symbiotic commensals (see Box 4.1) that grow on the surfaces of other plants and are common in most tropical rainforests. Numerous bromeliads (Bromeliaceae) can be seen on the trunks of several of the trees in this forest in Puerto Rico. (Photograph courtesy of J. Thomson.)

384 Chapter 19
pared by early European explorers to a dimly lit, soaring green cathedral with an expansive space reaching to a high, vaulted roof. “Jungles” are the consequences of disturbed, high-light environments on the margins of the forest, along roads, rivers, and in gaps, or where logging has thinned the canopy.
Because of this vertical light distribution within the rainforest, much of its productivity is concentrated high in the canopy. Plant life forms that are not present in most temperate forests greatly contribute to the diversity of tropical forests. These plants include many species of lianas (woody vines rooted in the ground) and epiphytes (plants that grow on other plants but are not rooted in the ground; see Figure 2.16), including many species of ferns, orchids, and bromeliads. Lianas and epiphytes are common in tropical forests because of the very low light levels near the ground. An alternative strategy to being a tree is to grow high up on a tree to gain access to light.
Most epiphytes and lianas have commensal relationships (the host neither benefits nor is harmed) with the trees on which they grow unless the burden of epiphytes and lianas becomes too large, weakening the tree and possibly pulling it down. Some species are not so benign, however. Strangler figs (Ficus spp., Moraceae) begin as a number of separate individual plants that send roots downward to the forest floor, develop thick trunks and canopies, and gradually envelop and kill the host tree (see Figure 10.7). The separate individual plants comprising the strangler ultimately merge to become a functional individual tree. Thus, this plant has a combined strategy of beginning life high in the canopy as an epiphyte, becoming rooted in the ground, which allows more growth, and eventually becoming a tree and outcompeting other species for light.
Moist tropical forests typically have highly infertile soils. In tropical rainforests, warmth and abundant moisture contribute to an extraordinary amount of respiration in the soil—not only by roots, but also by many other kinds of soil dwellers, particularly decomposing organisms. When CO2 is dissolved in water, it produces carbonic acid (that is why natural rainwater is always slightly acidic). The huge amount of soil respiration in tropical rainforests releases large amounts of CO2, much of which goes into solution in the soil. The net effect is that nutrient ions, which bind to soil (especially clay) particles less tightly than hydrogen ions bind to soil particles, tend to enter the soil solution. As a result, they are either taken up almost instantaneously by plants or washed away by the high levels of rainfall (see Chapter 4).
If the soils are so poor, how is it that tropical forests are so productive? Factors contributing to their high productivity are year-round water availability, warm temperatures, and duration of daylight varying little
PAGE PROOF: 2ND PASS
throughout the year. As for the mineral nutrients needed for growth, there are large quantities of nutrients in these forests, but they are mainly found in the living biomass rather than in the soil. Because decomposition is extremely rapid in these environments, nutrients cycle very rapidly. Like plants in many areas where nutrients are limiting, those in tropical forests are very good at acquiring and retaining nutrients. Particularly important in this regard are mycorrhizal associations (see Chapter 4).
Not only is a large proportion of the world’s biodiversity located in the Tropics; a large proportion of its human population is located there as well. The greatest growth in human populations is also found in the Tropics, along with poverty and diseases associated with poverty and poor living conditions. All of these factors can drive the rapid obliteration of natural communities by people. Although until recently much of the human population in the Tropics was rural, it is becoming increasingly urban. Lagos, Nigeria, for example, with about 13 million people, is one of the fastest-growing cities in the world. Unfortunately, this urbanization does not mean that there is less pressure on natural habitats; in many cases, the opposite is true. Economic, population, and political pressures have contributed to the destruction of a large proportion of the world’s rainforests (Table 19.3; see Chapter 22). Exploitation comes not only from within the tropical countries themselves, but also in great measure from individuals and corporations based in the United States, Europe, and Japan. Today all tropical rainforests are threatened by logging, and even more by the clearing of land for a range of purposes, including subsistence slash-and-burn agriculture and corporate-owned cattle ranching (see Chapter 22).
Logging and the clearing of land for cattle grazing or human habitation can have numerous effects on rainforests. Because most of the nutrients are tied up in biomass, logging, burning, and agriculture remove substantial amounts of nutrients from the system. The remaining soils are often very nutrient-poor, and they can quickly achieve a cementlike texture or become severely eroded. These changes can make agriculture impossible in a very short time—in other words, the purposes for which the forest was removed are not sus-
Table 19.3 Global tropical rainforest loss
|
Area (× |
106 km2) |
|
|
Region |
Original |
Remaining |
% lost |
|
|
|
|
|
|
Central/South America |
8.0 |
5.8 |
|
28 |
Asia/Pacific Islands |
4.4 |
2.2 |
|
39 |
Africa |
3.6 |
1.2 |
|
66 |
|
|
|
|
|
|
|
|
|
|

PAGE PROOF: 2ND PASS
tainable. Removal of rainforest from large enough areas can drastically affect regional rainfall levels. When the forest is present, rainfall is quickly taken up and transpired back into the atmosphere. When the forest trees are no longer there, the rainfall runs off, which leads to increasing erosion, particularly for forests on slopes. Even when the destruction is not so dramatic, as in selectively logged forests, non-native plant species can invade disturbed forests, threatening the regeneration of native species.
While habitat destruction is the key factor threatening tropical biodiversity, other human activities, from the illicit trade in rare species such as orchids and parrots brought into the United States and Europe, to the hunting of chimpanzees and other primates for the delicacy called “bush meat” in African cities, also endanger animal and plant species within these forests. These forms of forest destruction are not particularly new; wealthy nineteenth-century orchid collectors in England and Germany destroyed entire tropical forests to assure that their competitors could not collect orchids from them. However, the scale of recent damage dwarfs that of the past, and, of course, the destruction is cumulative.
Tropical Montane Forest
Tropical montane forests—also called elfin forests or cloud forests at the highest elevations—are higher-ele- vation neighbors of rainforests (Figure 19.4). They are called cloud forests because they are often swathed in clouds, and much of the moisture available to plants comes from deposition of condensation. They are cooler than tropical rain forests because they are at higher elevations. The trees are typically shorter, and there are many more epiphytes festooning the branches of trees.
Biomes 385
Overall diversity is lower than in rainforests, however, because there are fewer lianas. These forests are found at middle elevations in the mountains of Africa, South America, Central America, and New Guinea.
Seasonal Tropical Forests and Woodlands
When people think about the ecology of tropical regions, they typically think about rainforests, especially when they think about species diversity and conservation. However, many tropical regions are considerably drier than rainforests, and many of these forests are also rich in species. The seasonal tropical forest biome, with its associated species, is being lost very rapidly. In fact, these forests are disappearing—or have already disap- peared—even more rapidly than tropical rainforests, as they are often on soils that are much better for farming. This biome includes a range of types that grade from deciduous forest to thorn forest and woodlands.
Several geographic factors determine the locations of dry tropical forests and woodlands. First, they tend to occur relatively close to the Tropics of Cancer and Capricorn, rather then near the equator. This pattern is a result of the shifting throughout the year of the intertropical convergence—the latitude at which the trade winds from the Northern and Southern Hemispheres tend to converge (see Figure 18.7), causing a large amount of rainfall. The intertropical convergence is usually near each of these latitudes only once a year, whereas it passes over the equator twice a year. As a result, rainfall is highly seasonal, whereas closer to the equator, rainfall is more evenly spread throughout the year (see Figure 18.15). Second, dry forests and woodlands tend to occur in the rain shadows of mountain
Figure 19.4
A tropical montane forest in the Santa Elena Cloud Forest Reserve, Costa Rica. (Photograph © G. Dimijian/Photo Researchers, Inc.)
386 Chapter 19
Figure 19.5
The top view of a tropical deciduous forest in Palo Verde National Park, Costa Rica was taken during the rainy season; the bottom photograph is the same view shown in the dry season, when many of the trees are leafless. (Photograph by Donald L. Stone.)
ranges (see Figure 18.9) and on the western slopes of continents. For example, the eastern (Caribbean) side of the mountain range in Costa Rica is tropical rainforest, whereas the western (Pacific Ocean) side is dry tropical forest.
Tropical Deciduous Forest
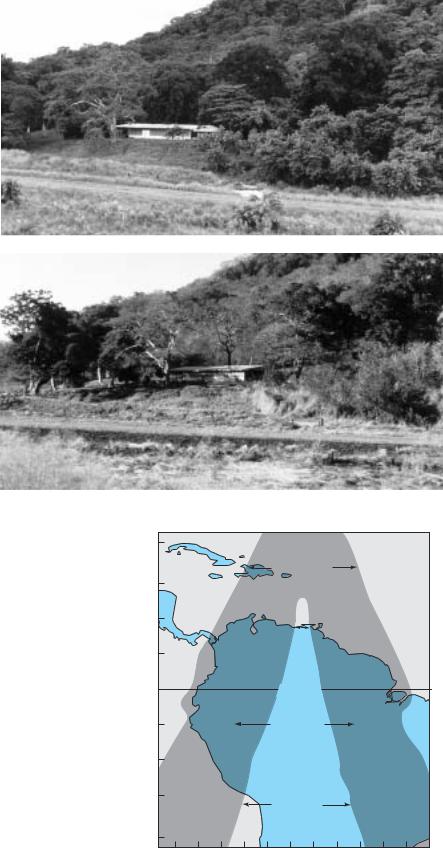
Tropical deciduous forests, also called tropical seasonal forests, include a range of community types, from wholly deciduous to semi-evergreen (Figure 19.5). The trees are drought-deciduous, meaning that they lose their leaves during the dry season. These communities are found in tropical areas that have pronounced wet and dry periods, and were once especially extensive in India and Southeast Asia, but also occurred in Central and South America and elsewhere. Depending
on the latitude, there might be a single wet and a single dry period, or two of each (Figure 19.6). Although most photosynthetic activity and plant growth occurs during the wet season, the dry season is often when flowering takes place. There may be advantages to flowering in the dry season for both animal-pollinated and wind-polli- nated plants. The lack of leaves makes the flowers more conspicuous to pollinators, and the pollen and nectar rewards may be one of the few food sources available to them. Similarly, wind-dispersed pollen can travel with fewer obstructions.
Tropical deciduous forests are similar in overall form and structure to rainforests, with a species richness lower than but approaching that of tropical rainforests in some
Figure 19.6
The length and number of rainy seasons depends on latitude. Near the equator there are two wet and dry seasons each year, while farther north and south there is a single rainy season in the summer and a dry season in the winter. The timing of the seasons is reversed north and south of the equator.
PAGE PROOF: 2ND PASS
23°N |
|
|
|
Dry season |
Single rainy |
Dry season |
|
season |
|||
15 |
|
||
|
|
||
10 |
|
|
|
|
Short |
|
|
5 |
dry |
|
|
season |
|
||
|
|
||
0 |
Equator |
|
|
5 |
Two rainy |
|
|
|
seasons |
|
|
10 |
|
|
|
15 |
Long dry |
|
|
|
|
||
|
season |
|
|
23°S |
|
|
|
J F M A M J J A S O N D |
|||
|
Month |
|
|

PAGE PROOF: 2ND PASS
areas. Localities with lower amounts of rainfall generally have shorter canopies and fewer species. Human populations are often very high in these biomes, and they face many of the same issues and problems as tropical rainforests.
Thorn Forest
Tropical areas that are even drier support thorn forests (also called thornwoods), named for the menacing armament of many of the plant species there (Figure 19.7). They are common from South America to Mexico, including those on limestone soils in the West Indies and
Biomes 387
Figure 19.7
A thorn forest on the island of Madagascar, off the east coast of Africa. This photo was taken after heavy rains in December. From August through November—the dry sea- son—the trees are parched and brown. (Photograph by N. Garbutt/ Indri Images.)
Central America, and in Asia (Burma, India, and Thailand), continental Africa, and Madagascar. They have very pronounced wet and dry seasons, and the total yearly rainfall may be half that of tropical deciduous forests. Species of Acacia and other woody members of this part of the pea family (Fabaceae) often dominate these communities. These plants have leaves that are divided into many
small leaflets, which helps to minimize water loss. Diversity is considerably lower than in tropical rainforests and deciduous forests, but can still be quite high compared with that in temperate systems. As in tropical seasonal forests, the trees are typically drought-deciduous. Succulents are also common, especially in drier areas. As the climate becomes even drier, these communities grade into either desert scrublands or tropical savannas.
Tropical Woodland
Forests are dominated by trees whose canopies touch each other. In contrast, the trees in woodlands are spaced apart, although the density of those trees can vary from closely spaced to sparse. Tropical woodlands are found in areas with strongly seasonal climates, like tropical deciduous forests, but which are drier or have less favorable soils. They occupy areas of Brazil (Figure 19.8) and interior southern Africa (miombo), and also occur in northern South America, the West Indies, northern Australia, and Burma and other parts
of Southeast Asia.
Figure 19.8
The vegetation on Brazil’s Sete Cidades National Park is an example of tropical woodland. (Photograph © J. Jangoux/Photo Researchers, Inc.)

388 Chapter 19
The trees in tropical woodlands range in height from 3 to 7 m. Most trees and shrubs have large, tough, semievergreen leaves and thick, fire-adapted bark. Many of the trees are members of the Fabaceae, as in thorn forests. Palms are also common, whereas succulents and spiny plants are rare. African woodlands—as well as savan- nas—are rich in large mammal species, including herds of elephants, giraffes, wildebeest, and their predators, such as lions and hyenas.
Temperate Deciduous Forest
Temperate deciduous forests are found in three disjunct areas of the Northern Hemisphere: eastern North America, eastern Asia, and Europe (Figure 19.9). These forests grow on young soils formed since the most recent glaciation; the lack of such forests in the Southern Hemisphere is probably a result of the absence of such soils there. Temperate deciduous forests are found in areas with continental or maritime climates with warm, moist summers and cold, sometimes snowy winters, with a range of 50–250 cm rainfall annually, distributed through the year. Diversity and productivity are generally moderate in these forests, but span a fairly wide range. In North America and Asia, this biome is known for the brilliant red, orange, and gold colors of its leaves in autumn, before they are shed for the winter. The dominant trees are 18–30 m tall. Growing seasons range from very short in the northernmost and high-altitude parts of this biome to very long in its southern portions. The genera of the dominant trees common to temperate deciduous forests on all three continents include Quercus (oak), Acer (maple), Fagus (beech), Castanea (chestnut), Ulmus (elm),
PAGE PROOF: 2ND PASS
Tilia (basswood or linden), Juglans (walnut), and Liquidambar (sweet gum). Different species of these genera occur on each continent. The diversity of local forest types on all continents may be very great, depending on the details of topography, climate, soils, and disturbance regimes. In North America, for instance, the eastern deciduous forest consists of many different forest subtypes (Braun 1950; Greller 1988).
At higher latitudes and elevations, temperate deciduous forest sometimes grades into mixed deciduousevergreen forest. An example is the hemlock-white pinenorthern hardwood forest in the northern United States (especially around the Great Lakes) and extending south in the upper elevations of the Appalachian Mountains. The dominant trees in this forest are a mixture of evergreen needle-leaved and deciduous broad-leaved species. Elsewhere, patches of pine-dominated communities can be found where the soil is especially shallow or infertile or there is a high frequency of fire. If such areas are large enough, they may be separately classified as temperate evergreen forest.
Tree species diversity in temperate deciduous forests is generally moderate, with 5–30 dominant tree species in any one place. However, the temperate deciduous forests of the southeastern United States are a center of plant diversity, with the highest tree species diversity in North America as well as a very high diversity of ferns and deciduous and shrubby angiosperms. For example, there are over 120 species of trees and over 3000 plant species in the southern Appalachian region. These forests escaped glaciation, and some areas have been continuously vegetated for as long as 200 million years. The dominant genus here is Quercus (oak, Fagaceae).
Other common canopy trees include numerous species of Carya (hickory, Juglandaceae), Acer (maple, Sapindaceae), Aesculus (buckeye or horse chestnut, Sapindaceae), and Magnolia (Magnoliaceae), as well as Tilia americana (basswood, Tiliaceae), Liriodendron tulipifera (tulip poplar, Magnoliaceae), and Betula lenta (black birch, Betulaceae).
Common in the understory are trees such as Cornus florida
Figure 19.9
A temperate deciduous forest in Michigan, here dominated by American beech (Fagus grandifolia, Fagaceae) and sugar maple (Acer Saccharum, Sapindaceae). (Photograph by S. Scheiner.)