

PAGE PROOF: 2ND PASS
C H A P T E R 3 Water Relations and Energy Balance
Plants, like everything else alive, need water to live, grow, and reproduce. Like animals, plants use water as the medium in which all of their biochemical reactions occur; unlike terrestrial animals, most plants also rely on water to a greater or lesser extent for support of their nonwoody parts and
for maintaining their physical structure. Much of the evolutionary history of plants on land is the story of increasing success in obtaining water, moving it to parts of the plant far from the source of the water, and being able to reproduce in a very dry atmosphere. We briefly review this history, pointing out some of the essential structures that evolved during the course of the history of land plants.
Terrestrial plants not only must obtain water, but are also faced with the problem of restricting their water losses. Intimately related to this problem is the challenge of maintaining their leaf temperatures within an acceptable range. In this chapter, we examine the evolutionary “solutions” to these interrelated problems and take a look at the physics of how plants exchange water and energy with their environments.
Adapting to Life on Land
Photosynthetic organisms originated in marine environments, where they were continuously bathed by water in which mineral nutrients were dissolved. The ancestors of terrestrial land plants were single-celled aquatic organisms that depended on having each cell in direct contact with water, obtaining nutrients from and being kept moist by the water in their immediate surroundings. Like modern photosynthetic aquatic organisms, these ancestors of terrestrial plants also depended on water for sexual reproduction, releasing their gametes into the water.
Early land plants did not differ dramatically from their aquatic ancestors in many of these characteristics. One of the earliest adaptations of the first land plants was the evolution of a cuticle, a waxy, nonliving covering over the exposed epidermal cells that prevents desiccation by the air. In bryophytes such as mosses, for example, water moves by diffusion from cell to cell. There are no specialized organs to take up water, nor to transport it to distant parts of the plant. The process is slow and not very efficient; as a consequence, none of the cells in a moss plant can get very far away from the substrate (soil or rock, for instance) that provides the water, so mosses are constrained to be very short.
42 Chapter 3
The dominant generation in bryophytes, including mosses, is the haploid gametophyte (see Chapter 8). Male gametophytes produce male gametes, which must swim to the female gametes. Because these plants are terrestrial, this is possible only when the moss shoots are covered with a film of water from rainfall or dew. The sperm swim along this film of water from a male plant to a female plant, eventually making their way to an archegonium containing an egg. This will work successfully only under certain restricted weather conditions, and males and females must grow close to each other.
Early vascular plants, which branched off from the ancestors of bryophytes in the Paleozoic era (see Chapter 21), had an enormous advantage over bryophytes for living in a terrestrial environment: their vascular systems allowed the transport of water much more rapidly and efficiently throughout the plant. Because they had evolved vascular tissues, they were able to develop specialized structures to take up water from the soil (roots) and woody tissue capable of supporting a trunk and crown. Without vascular tissue to take up water and conduct it over long distances, such structures could not exist. The early vascular plants also evolved a dominant sporophyte (diploid) generation. As the sporophyte became larger and longer-lived over the course of evolution, the gametophyte became smaller and shorterlived. However, the plants in these groups still depended on motile sperm to swim from the male to the female gametophyte, and had not evolved seeds.
The seed plants—seed ferns (now extinct), gymnosperms, and angiosperms—represent the greatest advances in the adaptation of plant life to a dry environment. The tissue that conducts water in this group of plants, the xylem, is capable of moving large volumes of water efficiently over long distances (Figure 3.1). Particularly in the angiosperms—the flowering plants— there is a great diversity of structure and physiological function related to the ability to withstand dry conditions.
The evolution of pollination removes many restrictions on the ability to reproduce in the dry conditions that dominate terrestrial life, including the requirement that male and female gametophytes be located adjacent to each other and that reproduction occur only under a very limited set of weather conditions.
The evolution of seeds was another key innovation. In non-seed vascular plants, such as ferns, dispersal occurs as haploid spores, produced by the diploid sporophyte plant. Both seeds and spores can disperse from the parent plant and can survive various environmental hazards, including dessication. However, seeds include a number of tissues, such as the seed coat, that allow for the detection of subtle variation in the environment (such as changes in daylength and temperature) and for
PAGE PROOF: 2ND PASS
the hormonal regulation of germination in response to environmental conditions, as well as for maternal provisioning of food for the embryo. Seed plants were, therefore, able to evolve far greater variation in dormancy and in seed provisioning than their non-seed ancestors.
Water Potential
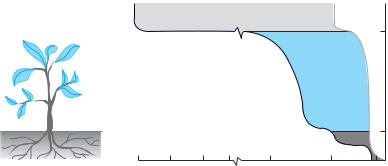
The ability of a plant to acquire water, the ability to move it to all cells throughout the plant, and the propensity of the plant to lose that water all depend on the water potential of the various parts of the plant and its immediate environment (Figure 3.2). One way of thinking about water potential is that it represents the difference in potential energy between pure water (which is defined as having a water potential of zero) and the water in some system, such as that in a plant cell or in the soil.
When water is bound up in some way, or has materials dissolved in it, the potential energy it possesses is less than that of pure water, and its water potential is negative. Analyzing the movement of water in plants in terms of water potential puts the study of plant water relations into the unifying context of thermodynamics. It makes it possible to study the movement of water in soils, in plants, and between plants and the atmosphere using the same concepts, terms, and units.
Water potential (Ψ ) in plants and soils can be broken down into four major components,
Ψ = Ψ π + Ψ P + Ψ m + Ψ g
where Ψ π is the osmotic potential, Ψ P is the pressure potential, Ψ m is the matric potential, and Ψ g the gravitational potential of the water in the system.
The osmotic potential is the component of water potential that is due to solutes dissolved in the water. It results from the difference in potential energy between pure water and water containing dissolved substances. Osmotic potential is either zero (in pure water) or negative, because the solutes reduce the capacity of the water to do work. The osmotic potential is capable of acting as a driving force for water movement such as when the movement of the solutes is restricted by a semipermeable membrane that allows water, but not solute molecules, to pass through it. The osmotic potential is the major component of water potential in living cells (such as those in roots and leaves), and thus is the major driving force by which water moves into those cells.
The pressure potential is a second key component of plant water potential, and is important both in living cells and in functioning xylem tissue (which is nonliving at maturity). It can be negative, zero, or positive in value, and is a function of the hydrostatic or pneumatic pressure in the system. When water is enclosed by

PAGE PROOF: 2ND PASS
Water Relations and Energy Balance 43
Simple perforation plate
Scalariform perforation plate
(A)
(D)
Simple perforation plate |
(E) |
|
Bordered pit
(B) |
(C) |
(F) |
|
Figure 3.1
Water-conducting xylem cells from the wood of a gymnosperm and several angiosperm trees. (A) A tracheid of Pinus lambertiana (sugar pine, Pinaceae), bent in several places to show its full length. Notice that the ends of the tracheid are not open. (B) An enlarged view of the central part of the tracheid in (A), showing pits in the side walls. (C) Xylem vessel element of Liriodendron tulipifera (tulip tree, Magnoliaceae), showing openings in the end wall (the scalariform perforation plate). (D) Xylem vessel element of
something that restricts its volume, such as a healthy plant cell enclosed by cell walls, the water presses on the walls, generating a positive pressure potential. This is the ordinary situation in a fully hydrated, or turgid,
Fagus grandifolia (beech, Fagaceae), with an open end wall (simple perforation plate). (E) Xylem vessel element of Populus trichocarpa (black cottonwood, Salicaceae). (F) Xylem vessel element of Ailanthus altissima (tree-of-heaven, Simarubaceae). Moving from A and B to E, the elements become progressively shorter, wider, and have larger openings in the end walls, all contributing to facilitating the passage of greater volumes of water more rapidly. Scale bars are all 100m. (From Carpenter and Leney 1952.)
plant cell under normal conditions, in which the action of the water is similar to that of air inflating a balloon. The opposite situation occurs when water is pulled through an open system, such as a garden hose or a
44 Chapter 3
PAGE PROOF: 2ND PASS
Atmosphere
Plant
Soil
–150 –100 –50 –10 |
–4 |
–2 |
0 |
Water potential (MPa) (Note change of scale)
Figure 3.2
Typical values (in MPa) for water potentials in the soil, in a plant, and in the atmosphere in a mesic environment. The curve at the left side of the shaded region indicates the values for atmosphere, plant, and soil water potentials when the atmospheric relative humidity is low and the soil is dry. Under these conditions, soil water potentials range from close to 0 at deeper layers to about –1.8 MPa near the soil surface, plant water potentials range from about –2.0 to about –4.0 MPa, and atmospheric water potentials range from about –5.0 to –150.0 MPa. The light gray curve at the right side of the shaded region indicates the values for moist soils and high atmospheric relative humidity; under these conditions, values for soil, plant, and atmosphere would be about 0 MPa, 0–0.75 MPa, and –1.0 to < –2.0 MPa respectively. Intermediate conditions would result in values falling within the shaded region. (After Etherington 1982.)
plant xylem vessel. In that case, the pressure potential is negative, and actually pulls the walls of the vessel inward. Both positive and negative pressure potentials exist in plants, and both are important driving forces for the movement of water within the plant and between the soil, the plant, and the atmosphere.
Gravitational potential results from the pull of gravity on water, and it is largely responsible for water draining from the largest pores of the soil in the first few days after a saturating rain (see Chapter 4). It is negative when the water is moving downward, since the water is losing potential energy. Matric potential is a result of the cohesive force that binds water to physical objects, such as cell walls and soil particles; it is always negative because water bound to an object has less potential energy than free water. The matric and gravitational potentials are generally much more important in soils than in plants.
The Soil-Plant-Atmosphere Continuum
Water always moves from larger to smaller values of Ψ . In plants and in the soil, this usually means moving from a region of less negative water potential to one of more negative water potential. The gradient in water
potential acts as the driving force for water movement from the soil, into and through the plant, and out to the atmosphere through transpiration.
The energy for moving water is provided without the expenditure of any energy by the plant itself. Energy is needed to lift the water up from the soil to the top of the plant, and to change the less energetic liquid water molecules inside of the plant into the more energetic molecules of water vapor in the atmosphere. The movement of water is driven by the gradient in water potential along this pathway, and the energy comes directly from the sun. In this way, plants are able to move vast amounts of water, sometimes over tremendous distances (in large trees), at no direct energy cost to themselves.
In a typical plant growing in moist soil and transpiring on a warm day, the values for water potential and its components might look like those in Figure 3.3A. Water moves from the soil into the plant roots, which have more negative Ψ , due primarily to the osmotic potential of the root cells. Then the water is drawn into the xylem vessels, which have still more negative Ψ values, due in this case to the negative pressure potential. Water moves into the leaves from the smallest veins, and out into the substomatal cavity (Figure 3.4). From there, when the stomata are open, water moves out into the atmosphere. Even in humid air (Figure 3.3B), the atmosphere surrounding the leaf has a highly negative water potential. The liquid water molecules ultimately gain the energy needed to move into a vapor state from the energy in sunlight, and the water throughout the rest of the pathway is pulled upward by the transpirational stream escaping from the leaves of the plant through the stomata.
Transpiration and the Control
of Water Loss
Plants transpire vastly more water than they use directly for plant functions. A large rainforest tree with a plentiful water supply might transpire as much as a thousand liters of water in a day (weighing 1000 kg—a ton of water lifted from the soil, transported up the trunk,

PAGE PROOF: 2ND PASS
Water Relations and Energy Balance 45
(A) |
Air (50% relative humidity): –9.4 MPa |
(B) |
|
Leaves: |
–2.3 MPa |
|
–0.5 to –2.5 MPa |
|
–2.3 |
–1.9 MPa |
–2.3 |
Xylem sap: |
|
|
–0.5 to –1.5 MPa |
|
|
Roots: –0.2 to – 0.4 MPa |
|
–2.3 |
–2.3 |
|
|
Moist soil: –0.1 MPa |
|
|
Figure 3.3 |
|
|
(A) Typical values for water potential from roots to stem |
–1.5 |
|
to leaves and atmosphere in an herbaceous mesophytic plant, with moist soil and air at 50% relative humidity.
(B) Water potential values in different parts of a Sequoiadendron giganteum (giant sequoia, Cupressaceae) tree that is transpiring rapidly. Here, the soil is moist and the air humid. The needles on the periphery of the canopy have the lowest water potentials, and the trunk has a higher water potential at the base than high in the canopy. (After Mohr and Schopfer 1995.)
Bundle sheath |
Mesophyll |
Xylem |
|
|
100 m |
Phloem |
Figure 3.4
Cross section of a leaf of Hordeum vulgare (barley, Poaceae), showing the upper and lower epidermis, which are each one cell thick; photosynthetic mesophyll cells, and a vascular bundle with xylem and phloem surrounded by a single-layered bundle sheath. Stomata are shown by arrows, and the substomatal cavities are visible inside the leaf, above the guard cells of the stomata. (From Esau 1977.)
46 Chapter 3
and lost through the leaves each day!). Temperate deciduous trees may lose up to 140 liters of water, but a typical conifer loses only 30 liters in a day. While most plants transpire fairly modest amounts of water, in one study, Salix fragilis (brittle willow, Salicaceae) growing in moist soil lost a remarkable 463 liters per day (Cermak et al. 1984).
The loss of water through transpiration can be described by a flux equation, much like the diffusion of CO2 into a leaf during the process of photosynthesis:
E = gleaf(wv) × (Ci(wv) – Ca(vw))
where E is the transpiration rate in mmol H2O/m2/s. The term m2 refers to the surface area of the leaf, usually the area of one side of the blade, but including both upper and lower sides for leaves with stomata on both surfaces. The transpirational flux is driven by the difference between the concentration of water vapor inside the leaf, Ci(wv), and that in the external atmosphere surrounding the leaf, Ca(vw), expressed in mol/m3. Sometimes the vapor pressure of water is used instead of vapor concentration in this equation, and the difference between water vapor in the leaf and the air is called the vapor pressure deficit (measured in kilopascals, KPa). The vapor pressure deficit of water is related directly to the water vapor concentration difference. The
leaf conductance to water vapor, gleaf(wv), is a measure of how readily water vapor moves into or out of the
stomata, and is defined as the rate at which water vapor flows into the leaf for a given difference between ambient and interc0ellular concentration; it is expressed in mmol H2O/m2/s and is the inverse of leaf resistance. The leaf conductance for water vapor flux consists of the boundary layer conductance (ga), cuticular conductance (due to the waxy leaf cutical; gc), and stomatal conductance (gs) to water vapor. Boundary layer conductance to water vapor is generally high. The thickness of the boundary layer of air surrounding a leaf depends on leaf size and shape as well as on ambient conditions, so it is not under the immediate control of the leaf. It has little effect on impeding water vapor flux. Cuticular conductance is very small and varies little: only limited water vapor leaves the leaf through the cuticle. In contrast, stomatal conductance is highly variable and is under the control of the plant: it is high when the stomata are open, and very low when the stomata close. Stomata are therefore very important in controlling the amount of water lost by the plant through transpiration.
The loss of large volumes of water through transpiration does not have negative effects on the plant when soil water is freely available. Transpiration is what moves water, along with dissolved mineral nutrients, from the roots to the leaves without the need for the plant to expend energy. As the soil becomes drier, however, the plant faces a dilemma. As CO2 moves into the
PAGE PROOF: 2ND PASS
leaves through the stomata, water moves out through transpiration. A plant that keeps its stomata closed to restrict water loss eventually faces what is essentially starvation, as it can no longer take in CO2. Angiosperms have evolved a greater diversity of mechanisms for coping with this dilemma, and for living with drought, than any other group of plants.
Strategies for Coping
with Water Availability Conditions
Plants experience drought in a variety of ways, depending on both environmental conditions where they live and on their adaptations to those conditions. Drought may be brief or prolonged, mild or extreme, occasional and unpredictable or regular in its onset and duration. It may occur when conditions are otherwise favorable for growth, or it may occur when it is too cold for plants to grow.
Plants have evolved various means of coping with limited water availability. These evolutionary solutions, or sets of coordinated adaptive traits, are known as strategies. Ecologists have come up with different terms to categorize some of these sets of traits, but it is worth remembering that these categories are designated for convenience rather than describing fixed, rigidly defined entities. In reality, each species has a unique set of characters for coping with the conditions in its environment.
Mesophytes are plants that live in moderately moist soils and generally experience only an occasional, mild shortage of available water. Mesophytes typically transpire as long as soil Ψ is greater than about –1.5 MPa, although some need soils wetter than –1 MPa, while others may be able to continue withdrawing water from the soil at water potentials as low as –4 MPa. When the soil becomes drier, mesophytic plants close their stomata and wait until conditions improve. In a typical mesic (moist) habitat, that may be a few hours or a few days. If the drought extends for many days (a few weeks is all that most of them can tolerate), many mesophytes will begin to die. Most crops, ornamental plants, forest trees, common meadow and forest wildflowers, and other familiar plants are mesophytes. Agricultural crops are generally the least drought-tolerant, and need soils that do not dry out too much in order to grow well.
Aquatic plants grow in standing water, and hygrophytes are found in permanently moist soils. Plant ecologists and amateur naturalists alike have always been fascinated with the adaptations of xerophytes, plants that live in regions with frequent or extended drought. Halophytes are plants that live in saline soils and possess unique adaptations for tolerating salt, as well as some characteristics in common with xerophytes. Halophytes experience a dilemma similar to that of xerophytes in that plants in both dry and saline soils must be able to extract water from soils with very negative

PAGE PROOF: 2ND PASS
water potentials. Some xerophytes continue to transpire in soils that are phenomenally dry, as low as –6 MPa. The halophyte Atriplex polycarpa (desert saltbush, Amaranthaceae) can continue to grow with leaf water potentials as low as –5 MPa. Even under well-watered conditions, its leaf water potentials are still at –2 MPa—a condition that would be fatal to most mesophytes.
Adaptation to environmental conditions can occur by means of either a fixed phenotype (in physiology, morphology, or anatomy) or a plastic phenotype (one that allows an individual plant to change in response to changing conditions). One type of phenotypic plasticity is the ability to acclimate (acclimatize) to changes in environmental conditions, such as drought, with either permanent or reversible changes in anatomy, physiology, or morphology. Permanent changes might include the production of leaves with different structural characteristics; once formed, these leaves would retain their structure. Reversible changes might include changes in the form of enzymes that function optimally at different temperatures, or changes in the osmotic potential of leaf cells. The ability to acclimate is important for plants because they are not able to move when conditions change: they can neither step into the shade when it is hot nor migrate to a warmer climate when it is cold.
Whole-Plant Adaptations to Low Water Availability
A plant experiences its environment very differently than one might expect from a cursory look at average
Water Relations and Energy Balance 47
conditions. Many desert annuals, particularly the shortlived desert ephemerals, complete their entire aboveground lives during the brief rainy season characteristic of many deserts. These plants spend most of their lives as seeds, which are able to survive extended dry periods. The seeds of these plants germinate only after prolonged rains, and the plants photosynthesize rapidly, have high rates of transpirational water loss, grow rapidly, then set seed and die as the rainy season ends. By “living fast and dying young,” they experience the desert as an environment with plentiful water for most of their lives as green plants. This strategy has been called drought avoidance.
Many desert shrubs lose all or some of their leaves during the long dry season and grow new leaves after the rains begin again (Figure 3.5). These drought-decid- uous plants are also practicing a form of drought avoidance, minimizing their water loss through transpiration by reducing their leaf area when water is not available. Drought-deciduous trees, such as Ceiba pentandra (kapok tree, Bombacaceae), are also common in seasonal tropical forests, which also experience predictable dry periods alternating with rainy seasons.
Many herbaceous perennials in deserts, arid grasslands, and other xeric (dry) environments are likewise largely dormant during the dry season, when most of the living material is found below ground or at the ground surface. The aboveground parts of many grasses, for example, die back to the ground during the dry
Figure 3.5
Fouquieria splendens (ocotillo, Fouquieriaceae), a drought-deciduous desert plant, in Joshua Tree National Park, in the Mojave Desert of eastern California. This species may produce several sets of leaves during a single year. (Photographs courtesy of P. Curtis.)

48 Chapter 3
PAGE PROOF: 2ND PASS
season. The advantage of dormancy during the dry season is, of course, reduction of water loss, enhancing survival during the unfavorable dry season. Its major disadvantage is that there is a long period of time during which the plant cannot photosynthesize, grow, or reproduce. Drought-deciduous plants generally have fairly high maximum photosynthetic and growth rates during the time when they are active, which help to compensate for the time when they are dormant. Many of these plants also have thick roots and underground stems that are capable of storing food for long periods of time. These structures enable them to survive extended drought periods and allow rapid production of leaves and stems when conditions become favorable again.
Another way in which some species avoid drought is to grow only in the wettest places within a dry environment, such as in low spots where water temporarily collects after rains, or along temporary streams. Such plants typically lack adaptations for extreme aridity and the seedlings will die if their seeds germinate in drier microhabitats.
There are, however, plants that are able to live and grow under remarkably dry conditions. These droughttolerant species, or true xerophytes, use a variety of means to “tough it out” in the desert. From the whole plant to the cellular and molecular levels, the morphology, physiology, and anatomy of these plants are adapted for life in xeric habitats.
Root morphology can be adapted to dry environments in a variety of ways. Roots can be extensive and shallow, as they are in many cactus species, enabling the plant to take up water after a brief rain that saturates only the top layers of soil. Other species have roots that extend deep into the soil profile, where the soil is slow to dry out. Phreatophytes have roots that extend so deeply into the soil that they reach the water table, gaining relatively permanent access to water—the ultimate drought avoidance mechanism.
The ratio of dry mass invested in roots to the mass of aboveground tissue (leaves, stems, etc.), called the root-to-shoot ratio, varies widely among plants. Xerophytes typically have much higher root-to-shoot ratios than do mesophytes. Consequently, they are able to take up more water, and they lose less water through transpiration. The cost of a high root-to-shoot ratio may be a reduction in maximum growth rates, both because of a reduction in the total leaf area, and therefore the amount of photosynthetic tissue, and because of the metabolic cost of maintaining the large root mass.
The form of the whole plant may also be an adaptation to limited water availability. Succulents, such as cacti, have the capacity to store large amounts of water in their tissues. The succulent tissue may be located in leaves, stems, or other plant parts. Cacti use stems instead of leaves as their photosynthetic organs. As a result, they have reduced surface-to-volume ratios, decreasing the total surface area capable of water loss and increasing the volume that retains water. Other desert plants also have photosynthetic stems. For example, in the genus Parkinsonia (palo verde, Fabaceae; Figure 3.6) in the Sonoran
Figure 3.6
Parkinsonia microphyllum (foothill palo verde, Fabaceae) (left) has extremely small leaflets (right), which reduce water loss by transpiration. Even these leaves are lost during the dry season, but photosynthesis continues at low rates in the green bark. (Photographs by S. Scheiner.)

PAGE PROOF: 2ND PASS
Water Relations and Energy Balance 49
Desert retain their leaves only during the rainy season, but remain capable of photosynthesis at low rates all year through their green bark.
In parts of the world where the soil freezes in winter, it can be difficult for plants to obtain liquid water from the soil for many months. Plants in these environments can experience severe drought stress in winter. This is one reason why deciduous trees and shrubs in the northern temperate zone and further north lose their leaves in winter, and it is one of the factors that limits the distribution of broad-leaved evergreen species northward.
Physiological Adaptations
When a plant begins to experience a water deficit, a series of physiological events is triggered. Hormones are produced and travel throughout the plant, signaling the onset of various changes in plant function. Cell growth and most protein synthesis slows, then ceases. As the plant experiences more prolonged water deficits, the allocation of materials to roots and shoots is adjusted, the stomata begin to close, and photosynthesis is inhibited. Leaves may begin to wilt. In some species, older leaves are allowed to dry out and die, freeing up available water to preserve younger leaves for a longer period.
In some species, specific measures for osmoregulation are initiated under dry conditions. These plants synthesize certain soluble compounds (proline and other low-molecular-weight nitrogen compounds as well as soluble carbohydrates) during drought. The resulting increase in solutes lowers the osmotic potential of the cells, leading to an influx of water by osmosis, which prevents turgor loss and wilting. Osmoregulation is particularly important in allowing many halophytes growing in saline soils (where soil water has a negative water potential largely due to a negative osmotic potential caused by dissolved salts) to maintain a favorable water potential gradient. Other halophytes have the ability to excrete salt.
Some plants, known as “resurrection plants,” have highly unusual adaptations that allow them to survive extended complete desiccation (Figure 3.7). These plants include many lichens and some mosses, ferns, and other non-seed plants, as well as a number of angiosperms (in the Scrophulariaceae, Lamiaceae, Poaceae, and Liliaceae). They are found in various parts of the world, but are most diverse and abundant in southern Africa. All of these different plants survive cellular dehydration by a highly coordinated set of processes. Drought-stable proteins are synthesized, phospholipid-stabilizing carbohydrates are incorporated into the cell membranes, and the cytoplasm may become gelated. In the dehydrated state, metabolism is brought almost to a halt. Rehydration when water is again available also occurs in a highly coordinated manner in which the cell components are reconstructed step by step.
Figure 3.7
Selaginella lepidophylla (spike moss, Selaginellaceae) is a “resurrection plant”. During times of extended drought, the plant has developed adaptations that allow it to virtually stop its metabolism. Although the plant appears to be dead (top), the advent of rain brings it “miraculously” back to life (below). (Photographs © W. P. Armstrong.)
At the other end of the water availability spectrum, adaptation to flooding is critical for survival in some habitats (Figure 3.8). Plants may be subjected to variation in the depth to which they are submerged (from waterlogged soil to total submergence of the shoot) as well as in the frequency, season, and duration of exposure to flood conditions. Coastal species, such as those in salt marshes, and species found in floodplains and in riparian (river and stream) margins are predictably exposed to flooding, but the frequency and severity of flooding in many such locations have been increasing over time (see Chapter 22), and may surpass the toleration of these plants. Other habitats, including low-lying areas in forests, meadows, and fields, also expose plants to occasional submersion, particularly in sites with clay soils that do not drain readily after heavy rains.

PAGE PROOF: 2ND PASS
Anatomical and
Morphological Adaptations
A wide variety of anatomical and morphological adaptations enable plants to survive and grow under very dry or very wet conditions. Among the most important are variations in stomatal numbers, arrangement, size and behavior, which vary widely among plants. Stomata are the gateway for most of the water leaving a plant. Stomatal conductance varies directly with the pore width (size of the stomatal opening). Pore width is controlled by the guard cells on either side of the opening, which continually change shape, widening or narrowing the opening (Figure 3.9). The movement of the guard cells is controlled hormonally by the plant; it is not a simple physical consequence of the guard cells passively wilting as water is lost, as once was thought.
The guard cells open and close the stomata primarily in response to three factors: light, CO2 concentration, and water availability. The action of the guard cells differs among the three photosynthetic pathways: C3, C4, and CAM (see Chapter 2). Light directly causes the stomata of C3 and C4 plants to open. The partial pressure of CO2 in the intercellular leaf space has a major influence on the guard cells, signaling the stomata to close when it goes up and to open when it goes down. At night, when photosynthesis ceases in C3 and C4 plants, the CO2 concentration in the intercellular leaf space increases, and the stomata close. During the day, photosynthetic capture of CO2 reduces the partial pressure of CO2 in the leaf, and the stomata are induced to open. In CAM plants (see Chapter 2), the partial pressure of CO2 drops at night as it is captured by PEP carboxylase, and the stomata open. During the day, malate is decarboxylated (CO2 is removed) and accumulates in the leaf, signaling the stomata to close (see Figure 2.13).
Plant water relations are critical in determining the behavior of the stomata. Stomata respond to both leaf water potential and ambient humidity (Figure 3.10). A declining water potential in the leaf will override other factors, such as CO2 partial pressure, to close the stomata; clearly, preventing desiccation is of more immediate concern than is maintaining photosynthetic rate.
The stomata of different plants vary in their sensitivity to these three factors. Mesophytes, particularly