

PAGE PROOF: 2ND PASS
Belize
1950
Honduras
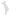 Nicaragua
Nicaragua
Guatemala
El Salvador |
Costa Rica |
Panama |
1970
1985
plant species may result from the increased incursion of edge effects into forests and other habitats.
Many habitats around the world are threatened by human activities. When these are rare or unique local habitats, reducing or destroying them may harm the only populations of a particular species in a region, or may harm the only remaining population of that species anywhere on Earth. Unique organisms and communities are found in many locations, and human encroachment is, in one way or another, threatening many of these places. Twenty-five biodiversity hotspots—threat- ened areas with very high species diversity—have been identified worldwide. These areas together contain 133,000 plant species, or about half of the world’s plant species (Figure 22.14, Table 22.1). The five most speciesrich of these hotspots support 20% of all plant species. Taken together, the 25 hotspots currently occupy 2.1 mil-
Global Change: Humans and Plants 457
Figure 22.13
Forest fragmentation in Central America between 1950 and 1985. Deforestation and fragmentation have increased in the years since 1985, although some forests are now protected in national parks, most notably in Costa Rica. Not only has the total area of intact forest declined dramatically, but the remaining forest fragments are small, often isolated from one another, and may experience large edge effects. Similar patterns are repeated for forests in many other parts of the world, and over a range of scales.
lion km2 (1.4% of Earth’s dry land surface); originally, these communities covered an area of 17.4 million km2. Together they have lost 88% of their original vegetation cover. Twenty of these hotspots are in tropical forests or mediterranean climate regions. Nine of them are on oceanic islands or archipelagos, where long isolation has led to the evolution of endemic species. Many of the others are isolated in other ways.
Not only rare species and rare communities are threatened, however. Less diverse but unique communities, sometimes with unusual organisms, including serpentine grasslands (see Box 15A), parts of the North American deserts, and some salt marshes, have also been subjected to various threats not only to their extent, but to their continued existence. In some cases, even originally very common species and ecosystems have been reduced to the point of becoming endangered, or have disappeared altogether. Climate change and human land uses have dramatically increased the extent of newly created depauperate deserts (with little vegetation of any kind, and with few uniquely adapted desert species) at the expense of fragile grasslands and shrublands. In the Sahel region of Africa, south of the Sahara, vast areas of once productive grasslands supporting large populations of people and animals have become shifting sands. In Lagan in central China (near the headwaters of the Yellow River), extensive grasslands have long supported humans and their grazing animals. They have now become barren wastelands supporting little vegetation, with recurrent sandstorms and vanishing rivers, lakes, and pastures, apparently due to a combination of climate change and intensive land use (particularly overgrazing as a result of human population pressure).
Tropical forests are among the best-known threatened biomes. About half of the species on Earth live in tropical rainforests, even though they now encompass only about 7% of terrestrial land cover. Many of these species are found only in very limited areas. For example, organisms in the seasonal tropical forests of the large island of Madagascar, off the east coast of southern Africa, have evolved in isolation for 121 million years, leading to some of the world’s highest levels of plant diversity and endemism. Madagascar has an estimated 10,000 plant species, of which about 80% are endemic.

458 Chapter 22
Table 22.1 “Hotspots” of biodiversity
PAGE PROOF: 2ND PASS
|
Original extent |
Remaining primary |
Area protected |
Number |
Number of |
|||
|
of primary |
vegetation (km2) |
(km2) |
of plant |
endemic species |
|||
Location |
vegetation (km2) |
(% of original extent) |
(% of hotspot) |
species |
(% of global plantsb ) |
|||
Tropical Andes |
1,258,000 |
314,500 |
(25.0) |
79,687 |
(25.3) |
45,000 |
20,000 |
(6.7%) |
Mesoamerica |
1,155,000 |
231,000 |
(20.0) |
138,437 |
(59.9) |
24,000 |
5,000 |
(1.7%) |
Caribbean |
263,500 |
29,840 |
(11.3) |
29,840 |
(100.0) |
12,000 |
5,000 |
(2.3%) |
Brazil’s Atlantic forest |
1,227,600 |
91,930 |
(7.5) |
33,084 |
(35.9) |
20,000 |
7,000 |
(2.7%) |
Choco/Darien/western |
260,600 |
63,000 |
(24.2) |
16,471 |
(26.1) |
9,000 |
2,250 |
(0.8%) |
Ecuador |
|
|
|
|
|
|
|
|
Brazil’s Cerrado |
1,783,200 |
356,630 |
(20.0) |
22,000 |
(6.2) |
10,000 |
4,400 |
(1.5%) |
Central Chile |
300,000 |
90,000 |
(30.0) |
9,167 |
(10.2) |
3,429 |
1,605 |
(0.5%) |
California Floristic Province |
324,000 |
80,000 |
(24.7) |
31,443 |
(39.3) |
4,426 |
2,125 |
(0.7%) |
Madagascara |
594,150 |
59,038 |
(9.9) |
11,548 |
(19.6) |
12,000 |
9,704 |
(3.2%) |
Eastern Arc and costal forests |
30,000 |
2,000 |
(6.7) |
2,000 |
(100.0) |
4,000 |
1,500 |
(0.5%) |
of Tanzania/Kenya |
|
|
|
|
|
|
|
|
Western African forests |
1,265,000 |
126,500 |
(10.0) |
20,324 |
(16.1) |
9,000 |
2,250 |
(0.8%) |
Cape Floristic Province |
74,000 |
18,000 |
(24.3) |
14,060 |
(78.1) |
8,200 |
5,682 |
(1.9%) |
Succulent Karoo |
112,000 |
30,000 |
(26.8) |
2,352 |
(7.8) |
4,849 |
1,940 |
(0.6%) |
Mediterranean basin |
2,362,000 |
110,000 |
(4.7) |
42,123 |
(38.3) |
25,000 |
13,000 |
(4.3%) |
Caucasus |
500,000 |
50,000 |
(10.0) |
14,050 |
(28.1) |
6,300 |
1,600 |
(0.5%) |
Sundaland |
1,600,000 |
125,000 |
(7.8) |
90,000 |
(72.0) |
25,000 |
15,000 |
(5.0%) |
Wallacea |
347,000 |
52,020 |
(15.0) |
20,415 |
(39.2) |
10,000 |
1,500 |
(0.5%) |
Philippines |
300,800 |
9,023 |
(3.0) |
3,910 |
(43.3) |
7,620 |
5,832 |
(1.9%) |
Indo-Burma |
2,060,000 |
100,000 |
(4.9) |
100,000 |
(100.0) |
13,500 |
7,000 |
(2.3%) |
South-central China |
800,000 |
64,000 |
(8.0) |
16,562 |
(25.9) |
12,000 |
3,500 |
(1.2%) |
Western Ghats/Sri Lanka |
182,500 |
12,450 |
(6.8) |
12,450 |
(100.0) |
4,780 |
2,180 |
(0.7%) |
Southwest Australia |
309,850 |
33,336 |
(10.8) |
33,336 |
(100.0) |
5,469 |
4,331 |
(1.4%) |
New Caledonia |
18,600 |
5,200 |
(28.0) |
526.7 |
(10.1) |
3,332 |
2,551 |
(0.9%) |
New Zealand |
270,500 |
59,400 |
(22.0) |
52,068 |
(87.7) |
2,300 |
1,865 |
(0.6%) |
Polynesia/Micronesia |
46,000 |
10,024 |
(21.8) |
4,913 |
(49.0) |
6,557 |
3,334 |
(1.1%) |
Totals |
17,444,300 |
2,122,891 |
(12.2) |
800,767 |
(37.7) |
* |
133,140 |
(44%) |
aMadagascar includes the nearby islands of Mauritius, Reunion, Seychelles, and Comoros. bThe total number of plant species globally is here estimated to be ~300,000.
*These totals cannot be summed due to overlapping between hotspots.
Madagascar also has some of the world’s most highly threatened habitats. Although forest destruction began many years ago, it accelerated greatly in the twentieth century. Between 1960 and 1990, slightly less than half the forested areas of Madagascar were destroyed. Today, less than 10% of the original (pre-eighteenth-century) forests remain.
Deforestation is occurring in tropical rainforest habitats worldwide, and is threatening or leading to the extinction of many species. Other forests, including highly diverse dry tropical forests, are also severely threatened. Climatic effects sometimes interact with other factors to intensify damage to tropical forests and other biomes. For instance, the 1997–1998 El Niño event caused uniquely ferocious and damaging fires in the rainforests of the island of Borneo, exacerbating the damage already done by extensive logging and land clear-
ing. Some scientists believe that global warming may have intensified these climatic effects.
It is difficult to get precise measurements of how much deforestation has already occurred, or of current regional rates of deforestation. Satellite imaging has greatly aided our ability to track deforestation, particularly in combination with new methods for image analysis. Estimated rates of deforestation differ among regions. Deforestation and logging have been accelerating in the Amazon in the 1990s (Figure 22.15). Similar rates of deforestation are occurring in tropical Africa (1.3–3.7 million ha/year: Boahene 1998). While the rainforests of Southeast Asia (Cambodia, Indonesia, Laos, Malaysia, Myanmar, Thailand, and Vietnam) cover a much smaller area in the decade from the mid-1970s to the mid-1980s, almost as much rainforest there was lost each year as in the Brazilian Amazon. Vast areas of

PAGE PROOF: 2ND PASS
Global Change: Humans and Plants 459
|
|
|
|
|
|
|
|
Philippines |
California |
|
|
Mediterranean |
Caucasus |
South-Central |
|||
|
|
|
|
China |
||||
|
|
Basin |
|
|
|
|||
Floristic |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Province |
|
Caribbean |
|
|
|
Indo-Burma |
|
|
|
Mesoamerica |
Brazil’s |
|
|
|
|
Polynesia/ |
|
|
|
|
|
|
Micronesia |
|||
|
Cerrado |
|
|
|
|
|||
|
|
|
|
|
|
Western |
||
|
|
Tropical |
Western |
|
|
|
||
|
|
|
|
|
|
|||
Polynesia/ |
Choco/ |
Andes |
|
African |
|
|
Ghats and |
|
Micronesia |
Darien/ |
|
|
Forests |
|
|
Sri Lanka |
Wallacea |
|
Western |
|
Brazil’s |
|
|
|
Sundaland |
New |
|
Ecuador |
Central |
Succulent |
|
|
|
Caledonia |
|
|
Atlantic |
|
|
Madagascar |
||||
|
|
Chile |
Karoo |
Eastern Arc |
|
Southwest |
||
|
|
Forest |
|
|||||
|
|
|
|
and Coastal |
|
|
Australia |
|
|
|
|
|
|
|
|
||
|
|
|
|
Cape Floristic |
|
|
|
|
|
|
|
|
Forests of |
|
|
New Zealand |
|
|
|
|
|
Province |
|
|
||
|
|
|
|
Tanzania/Kenya |
|
|||
|
|
|
|
|
|
|||
Figure 22.14
Twenty-five regions of the world identified as biodiversity hotspots have exceptional densities of species diversity and endemism. The hotspots are located within the colored regions, but may make up only parts of those regions. (After Myers et al. 2000.)
ancient forest were converted to agriculture or cut for timber. Similar rates of forest loss continue there to the present day. If current rates of deforestation continue, the world’s tropical forests will disappear within 100 years, existing only in tiny, isolated fragments (see Figure 22.13).
ha/yr) |
2.0 |
|
|
1.5 |
|
|
|
6 |
|
|
|
rate (10 |
1.0 |
|
|
Deforestation |
0.5 |
|
|
0.0 |
|
|
|
|
1992–94 |
1995–98 |
|
|
1990–91 |
Time period
Figure 22.15
Deforestation rates in the Brazilian Amazon have accelerated during the 1990s. Forest losses are expressed in millions of hectares a year. Deforestation rates in other parts of the Amazon basin have also been increasing. (Data from Laurance 2000b.)
Invasive Species and Other Threats to Biodiversity
Other major global threats to biodiversity include overexploitation of species by hunting, fishing, or collecting and the spread of invasive exotic species. Overexploitation more obviously threatens animals than plants, but plants are not immune to this threat. First, logging and removal for other reasons have greatly reduced the populations of some plant species. Several plant taxa, particularly orchids and cacti, are threatened by amateur collectors (Orlean 2000). While collecting and selling plants taken from natural populations is illegal in many areas, steep prices and minimal fines conspire to maximize these depredations. Second, many plant species are threatened indirectly by overexploitation of other species. Killing major predators, for example, can lead to large rises in herbivore populations, resulting in overgrazing or overbrowsing. The extensive loss of understory vegetation in forests all over the northeastern United States in recent years is due in large part to a tremendous rise in populations of white-tailed deer. The rise in deer populations is in turn the result of a number of factors, from suburbanization patterns that fragment forests and increase edge habitats, which favor the deer, to declines in predators and hunting (hunters are unwelcome in suburban backyards). The rise in deer numbers threatens forest regeneration because deer browsing kills tree saplings.
The spread of exotic and invasive species is increasingly threatening ecosystems all over the world (see Chapter 14). This phenomenon has serious consequences both for humans and for the preservation of

460 Chapter 22
native species and natural communities (Lodge 1993). Human activities such as deliberate and accidental species introductions, forest fragmentation, logging, and other disturbances have all been factors in the invasion and spread of non-native species. Dukes and Mooney (1999) raise the provocative suggestion that global climate change, including nitrogen deposition and rising atmospheric CO2, may preferentially enhance the success of invaders.
Exotic plants have invaded forests in many parts of the world; these species may have substantial effects on the retention and restoration of native species. New Zealand, for example, now has at least as many naturalized exotic plants as native species (Allan 1961; Webb et al. 1988). These introduced species have modified the vegetation of large areas in New Zealand. While native species have been not been driven to extinction there by the exotic species, the native plants appear to have been locally displaced and diminished in abundance by the invaders in many cases.
Grasslands, deserts, and other plant communities have also been greatly altered by invasive species. Invasive plant species can decrease the diversity of native plants and disrupt ecosystem processes, altering nitrogen cycling, fire regimes, and other fundamental characteristics of ecosystems (Higgins et al. 1999; Vitousek et al. 1996; Mack and D’Antonio 2001). In addition, biological invasions by exotic plants can exact enormous economic costs, including decreases in timber growth rates and forest value. These ecological and economic impacts can be so great that some ecologists have urged recognition of biological invasions as one of the most important forms of global change affecting ecosystems today.
Human Populations and Land Use Patterns
The number of people in the world has grown dramatically since the Industrial Revolution, and is continuing to grow rapidly (Figure 22.16). The consequences of global increases in the human population have been a cause for alarm, debate, and controversy for many years—as exemplified by the wide range of reactions to the much-disputed book by Paul Ehrlich, The Population Bomb (1968). Some of the predictions made regarding human population growth have been seriously flawed and alarmist, but attempts at outright dismissal of the concerns about its consequences have also been highly flawed. Much of the disagreement over and interest in the implications of human population growth has concerned the effects it might have on food supplies, and more generally on human societies. Less popular attention has focused on the possible environmental effects of growing human populations, although both environmentalists and social scientists have expressed concern about these effects.
PAGE PROOF: 2ND PASS
(billions) |
6.0 |
|
|
|
|
|
5.0 |
|
|
|
|
|
|
population |
|
|
|
|
|
|
3.0 |
|
|
|
|
|
|
|
4.0 |
|
|
|
|
|
human |
2.0 |
|
|
|
Revolution |
|
|
|
|
|
|
Industrial |
|
Global |
1.0 |
|
|
|
c. 1750 |
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
0 |
1 |
500 |
1000 |
1500 |
2000 |
|
|
|
|
Year |
|
|
Figure 22.16
Worldwide human population growth, estimated over the past 2000 years.
Recent growth in human populations has been much greater in developing than in industrialized regions, and tends to be highest in the Tropics, where a large proportion of the people are living at or below subsistence levels. Population growth has slowed in many of the developed, industrialized countries. As a result of these population growth patterns, increasingly greater proportions (and absolute numbers) of the world’s people live in developing countries, and an increasingly large proportion of the world’s people are children who live in poverty in those countries (Figure 22.17).
This rapid human population growth could have large environmental consequences (such as increased resource use, habitat destruction, and species extinctions) as well as economic and political effects and implications for global stability. But how much of the ongoing deforestation and other damage to natural communities can we actually ascribe to human population densities and population growth? How accurate is it to assume that increasing human populations directly affect forests or any other plant communities? Human societies are complex systems, and it is not surprising that human populations affect the environment in indirect and multifactorial ways. It is an oversimplification to assume that human population size is directly correlated with damage to the environment. Instead, both local and international economic and political forces interact with population size to influence changes in land use (Lambin et al. 2001).
Social scientists who seek to characterize and quantify the interactions between humans and their environments disagree about the extent to which different factors, including population size, are responsible for environmental problems such as deforestation. On the one hand, there is solid evidence that across a wide number of developing countries, population growth results

PAGE PROOF: 2ND PASS
Global Change: Humans and Plants 461
Age (years)
110 |
|
|
Industrialized |
|
|
|
|
|
|
|
|
|
|
|
|
Developing |
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
100 |
|
|
nations |
|
|
|
|
|
|
|
|
|
|
|
|
nations |
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
90 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
80 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
70 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
60 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
50 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
40 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
30 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
200 |
100 |
|
|
0 |
|
|
|
|
100 |
|
|
200 |
300 |
|
400 |
|
|
500 |
|
|
600 |
|||||||||||
|
|
|
Millions of people |
|
|
|
|
|
|
|
|
|
|
|
Millions of people |
|
|
|
|
|
|
|
|
|
|
|||||||
Figure 22.17
Global age pyramid. The total number of people, in millions, in each age category, from birth to 110 years, in industrialized and in developing nations. The total number of people in developing nations is far greater than in industrialized nations. The distribution of ages is also different, with relatively more children and more people at younger ages in the developing countries. The “baby boom” of people now in their thirties to early fifties is clearly evident as a bulge at those ages in the industrialized nations, but not in the developing nations. The number of very old people (over 90) is very small for both groups of nations; the maximum human age (not clearly visible here) is not different for the two groups. (Data from United Nations records.)
in greater deforestation (Ehrhardt-Martinez 1998). This has also been demonstrated at a global scale, although the negative relationship between human populations and declining forest area may have lessened in recent years, at least in some countries (Mather and Needle 2000).
In studies focused at the scale of particular countries or regions, there have been contradictory results. In two studies of the effects of human population density on deforestation in the Amazon basin of Brazil, its effects were at best indirect. Pfaff (1999) found that in the Brazilian Amazon, population density did not directly determine tropical forest clearing. Rather, many factors were implicated, from land characteristics and distance to markets to government development projects. However, Pfaff also found that while a static measure of population size was not a good predictor of deforestation, the first settlers in a region had a far greater effect on forest destruction than later immigrants. He concluded that the timing and distribution of population growth were more important than its absolute magnitude in determining its impact.
In the second study, Parayil and Tong (1998) found that population size and growth were not directly responsible for deforestation in the Amazon basin. Instead, multiple agents were involved, operating at different scales. Major agents responsible for deforestation included large-scale clearing of forests to create cattle ranches and logging of forests for lumber (both gener-
ally by large multinational corporations). The third major agent of forest destruction was the very smallscale clearing of forests for farming (slash-and-burn agriculture) by long-term residents as well as by impoverished immigrants to the region from other parts of the country. Thus, Parayil and Tong found that factors other than population growth were the major direct contributors to plant community destruction at a regional scale.
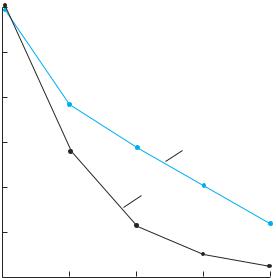
Patterns of forest loss and fragmentation are also changing (Laurance 2000b). In past decades, many of the largest-scale forest losses in the Amazon basin occurred at its southern and eastern edges. The development of major new highways, however, is now bringing settlers and loggers directly into the heart of the central Amazon forest, leading to great increases in fragmentation and forest loss (Figure 22.18; Laurance et al. 2001). This observation reinforces Pfaff’s argument that the distribution of human populations is at least as important as the absolute number of people in a country. The opening of new roads in forest areas also was found to be the most important factor in deforestation in the rainforest of Cameroon, in central Africa (Brown and Ekoko 2001). In that study, multiple agents (from the government to corporate loggers to local village interests and needs) and causes of deforestation acted synergistically to the detriment of the natural systems.
Patterns that are broadly similar in their complexity, but different in important details, have been found in human-environment interactions in the United States.
462 Chapter 22
|
30 |
|
|
|
|
|
|
25 |
|
|
|
|
|
deforested |
20 |
|
|
|
|
|
15 |
|
|
|
Paved highways |
||
Percent |
|
|
|
|
||
10 |
|
|
|
|
|
|
|
|
|
All roads |
|
||
|
|
|
|
|
||
|
|
|
|
and highways |
|
|
|
5 |
|
|
|
|
|
|
0 |
|
11–25 |
26 – 50 |
51– 75 |
76 –100 |
|
|
|||||
|
|
|||||
|
0 –10 |
|||||
Distance from road (km)
Figure 22.18
Effect of proximity to roads on deforestation in the Brazilian Amazon. The percentage of the original closed-canopy forest that was destroyed by 1992 is shown as related to distance from paved highways and from all roads and highways. Forests close to any roads, particularly paved highways, are far more likely to be lost than are those farther away. (After Laurance et al. 2001.)
In the past half-century, two distinct patterns of changing residential land use have had large effects on the reduction and fragmentation of wildlands. Both are, on the surface, seemingly innocuous, and stem in part from the desire of people to get out of the cities and live in closer contact with nature. The first of these is the rise of suburbanization—the establishment of vast low-densi- ty areas surrounding metropolitan centers, particularly in North America. The consequence has been a shift in population from cities to “metropolitan regions” sprawled over thousands of square kilometers. This shift has changed land that was once agricultural or occupied by natural communities into a mix of homes on large plots of land, with commercial development at some distance. The second, more recent change has been the incursion of vacation and retirement homes into wilderness areas, particularly in coastal, montane, and desert areas (Bartlett et al. 2000). While this land use does not consume as much land as suburban development, it can cause fragmentation of natural areas, and results in changes in fire regimes, erosion, and incursions of invasive plant and pest species.
Local planning and policy decisions that create suburban development patterns, in contrast to traditional towns or cities, have other unintended consequences in addition to the direct destruction of natural commu-
PAGE PROOF: 2ND PASS
nities and conversion to human-dominated landscapes. They lead directly to far greater per capita fuel consumption, because suburban residents have no choice but to commute long distances to work, to shop, to attend school, and for social and recreational opportunities. The most fuel-efficient vehicle driven by a commuter with a 100-mile round trip to work, or by a suburban parent who racks up 25,000 miles per year in everyday activities, will have far more impact than even a very large sports utility vehicle driven very few miles by a person living in a compact neighborhood where it is possible to walk or drive short distances for most activities. Recent efforts to build traditional towns and neighborhoods once again offer the opportunity to reverse some of these effects (Langdon 1997).
A Ray of Hope?
Is all gloom and doom? Is the world going to hell in a handbasket? There are clearly reasons to think so. However, there are also people working mightily to change the statement so often heard that “all this will happen if the current rates of [fill in your favorite bad thing] remain unchanged.” Perhaps some of these things can be changed. Human behavior is not immutable, and if current practices are altered, some of the consequences of our actions can also be changed.
Although there are many factors acting to continue our current momentum, it is certainly not impossible, for instance, to begin to bring carbon emissions in the United States into line with the kinds of reductions that have been accomplished in Western Europe. If we are concerned about the environmental damage being caused by unrestrained suburban sprawl in North America, there are workable, popular, and economically feasible alternatives to current environmentally costly land development patterns (for example, see Duany and Platt-Ziederbeck 2001). People are working in many countries to reduce carbon emissions, stop deforestation and habitat fragmentation, and restore damaged or destroyed ecosystems. We have the option of supporting efforts to protect species and habitats locally and around the world, and of changing our own habits and practices to reduce our personal impact on the environment. We also have the option of acting politically to alter current patterns of environmental destruction. The consequences of doing nothing should be obvious.
Summary
Earth is experiencing one of the most rapid periods of change in its history. The global carbon cycle is central to many of these changes. Anthropogenic increases in atmospheric CO2 and other greenhouse gases are expected to cause global climate changes that include warmer

PAGE PROOF: 2ND PASS
Global Change: Humans and Plants 463
temperatures, changes in precipitation patterns, and possibly changes in other climatic factors such as storm intensity and frequency. The increase in atmospheric greenhouse gasses is caused by a number of factors, chief among them fossil fuel combustion and deforestation. There is now strong evidence that many of the predicted climate changes are already occurring. Predictions for the future, taking current trends into account, are for warmer climates everywhere, with the greatest changes in polar regions. Rainfall patterns will shift, with some
areas becoming wetter and others drier, and both flooding and drought will increase.
Other anthropogenic global changes include habitat destruction and fragmentation, nitrogen deposition, and the spread of exotic invasive species. Nitrogen deposition and habitat decline and fragmentation will result in the decline or extinction of sensitive species. Together, these changes will result in major shifts in species’ distributions, changes in plant community composition and ecosystem productivity, and declining biodiversity.
Additional Readings
Classic References
Arrhenius, S. 1918. The Destinies of the Stars. G. P. Putnam’s Sons, New
York.
Hardin, G. 1968. The tragedy of the commons. Science 162:1243–1248.
Woodwell, G. M. 1970. Effects of pollution on the structure and physiology of ecosystems. Science 168: 429–433.
Contemporary Research
Aber, J. D. 1992. Nitrogen cycling and nitrogen saturation in temperate forest ecosystems. Trends Ecol. Evol. 7: 220–223.
Hughes, L. 2000. Biological consequences of global warming: is the signal already apparent? Trends Ecol. Evol. 15: 56–61.
Mack, R. N., D. Simberloff, W. Mark Lonsdale, H. Evans, M. Clout and F. A. Bazzaz. 2000. Biotic invasions: Causes, epidemiology, global consequences and control. Ecol. Appl. 10: 680–710.
Jefferies, R. L. and J. L. Maron. 1997. The embarrassment of riches: Atmospheric deposition of nitrogen and community and ecosystem processes.
Trends Ecol. Evol. 12: 74–78.
Additional Resources
Langdon, P. 1997. A Better Place to Live: Reshaping the American Suburb.
University of Massachusetts Press, Amherst.
Naeem, S., F. S. Chapin III, R. Costanza, P. R. Ehrlich, F. B. Golley, D. U. Hooper, J. H. Lawton, R. V. O’Neill, H. A. Mooney, O. E. Sala, A. J. Symstad and D. Tilman. 1999. Biodiversity and ecosystem functioning: Maintaining natural life support processes. Issues in Ecology No. 4. Ecological Society of America, Washington DC.
Vitousek, P. M, H. A. Mooney, J. Lubchenco and J. M. Melillo. 1997. Human domination of Earth’s ecosystems. Science 277: 494–499.