

Строение рнк
Отличия от ДНК:
по локализации (цитоплазма);
по функциям (обеспечивает биосинтез белка);
по размерам;
по строению (содержит У вместо Т, сахар — рибоза).
РНК бывает нескольких типов — иРНК, рРНК, тРНК, гяРНК (гетерогенная ядерная РНК), мяРНК (малая ядерная РНК).
Вторичная структура — всегда одна цепь (у тРНК — «лист клевера») (рис. 18.3).
Третичная структура — у тРНК формируется самостоятельно и похожа на объемную букву L; у рРНК и иРНК образуется в связи с белками (рРНК + белок = рибосома, иРНК + белок = информосома).
Тема 19. Обмен нуклеопротеинов
Н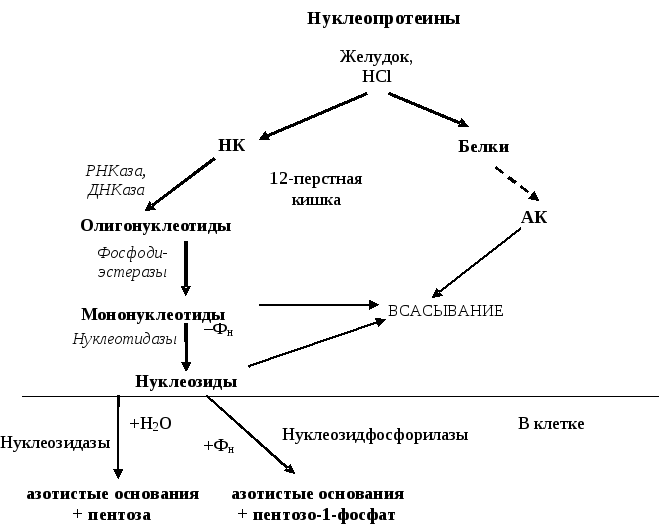 а
рисунке 19.1 представлена схема переваривания
и всасывания нуклеопротеинов в ЖКТ.
а
рисунке 19.1 представлена схема переваривания
и всасывания нуклеопротеинов в ЖКТ.
Рис. 19.1. Переваривание и всасывание нуклеопротеинов в ЖКТ
Как правило, экзогенные АО, нуклеозиды и нуклеотиды не используются в клетке для синтеза собственных нуклеиновых кислот. Они разрушаются до конечных продуктов и выводятся из организма.
Конечные продукты распада пиримидинов — β-аланин, β-аминоизомасляная кислота, NH3,CO2.
Конечный продукт распада пуринов — мочевая кислота (рис. 19.2).
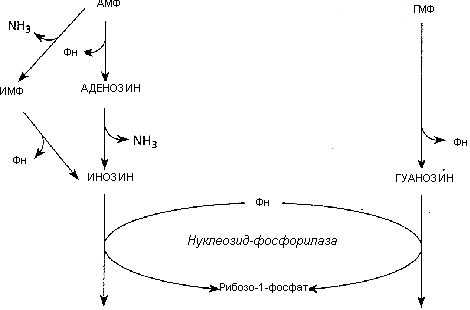
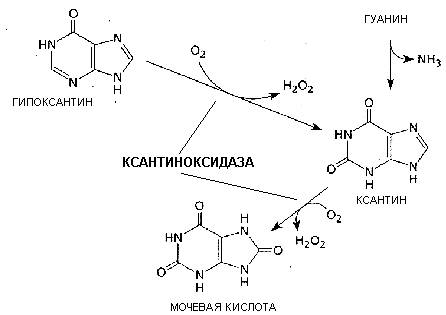
Рис. 19.2. Распад пуринов
Мочевая кислота содержит нерасщепленное пуриновое кольцо, поэтому плохо растворяется в воде. У человека мочевая кислота является конечным продуктом метаболизма и выводится с мочой.
Биосинтез нуклеотидов
Существует 2 пути биосинтеза нуклеотидов в клетке. Во-первых, путь повторного использования АО и нуклеозидов (не только экзогенных, но и образовавшихся в клетке в процессе репарации ДНК или при распаде «отработавших» РНК). Наиболее активно протекает в клетках интенсивно размножающихся тканей (эмбриональных, регенерирующих, эпителиальных, опухолевых). Во-вторых, синтез de novo (из низкомолекулярных предшественников).
Пути повторного использования АО и нуклеозидов: наличие этих путей позволяет использовать синтетические аналоги пуринов и пиримидинов для химиотерапии опухолей и лечения вирусных инфекций (например, 5-фторурацил, меркаптопурин, ацикловир, АЗТ и др.). Такие препараты включаются клеткой в состав нуклеотидов, встраиваются в молекулу ДНК и вызывают цитотоксический эффект.
АТФ АДФ

1
 .
Нуклеозид
Нуклеозидмонофосфат НТФ
.
Нуклеозид
Нуклеозидмонофосфат НТФ
Нуклеозидкиназа
Этот путь чаще используется для реутилизации пиримидинов (тимидинкиназа, цитидинкиназа).
2. Синтез нуклеотидов на основе готовых азотистых оснований больше характерен для пуринов и проходит в 2 этапа:
а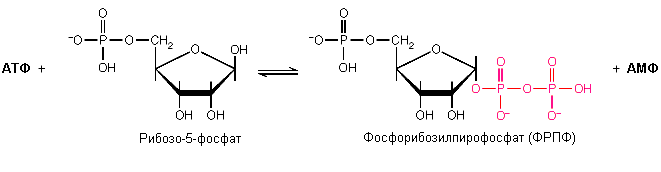 )
образование активной формы рибозо-5-фосфата
(фосфорибозилпирофосфата):
)
образование активной формы рибозо-5-фосфата
(фосфорибозилпирофосфата):
б) взаимодействие ФРПФ с азотистым основанием:
А О
О
 Нуклеозидмонофосфат НТФ
Нуклеозидмонофосфат НТФ
Фосфорибозил
трансфераза
De novo синтез пуриновых нуклеотидов
О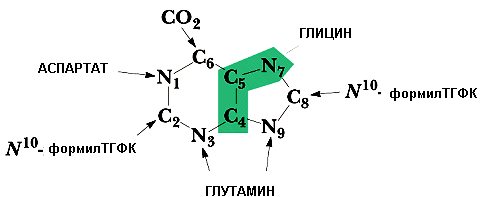 собенностью
синтеза пуриновde
novo(рис. 19.3) является
то, что за основу берется рибозо-5-фосфат;N-гликозидная связь
формируется уже на ранних этапах синтеза;
и только затем синтезируется пуриновое
кольцо.
собенностью
синтеза пуриновde
novo(рис. 19.3) является
то, что за основу берется рибозо-5-фосфат;N-гликозидная связь
формируется уже на ранних этапах синтеза;
и только затем синтезируется пуриновое
кольцо.
Источником всех атомов азота для пуринового ядра являются аминокислоты (глицин, глутамин, аспартат). Источники атомов углерода: СО2, метенил- и формил-ТГФК (ТГФК - активная форма фолиевой кислоты, В9). Общим предшественником для адениловых и гуаниловых нуклеотидов является инозинмонофосфат (ИМФ).
Р ибозо-5-фосфат
+ АТФ Фосфорибозилпирофосфат
+ АМФ
ибозо-5-фосфат
+ АТФ Фосфорибозилпирофосфат
+ АМФ
 ФРПФ-
(ФРПФ)
ФРПФ-
(ФРПФ)
синтетаза +глн
Амидотрансфераза