

Нервно-гуморальные факторы регуляции митоза
Среди факторов нервно-гуморальной регуляции митоза особое место отведено двум ключевым белкам: циклины и циклин-зависимые киназы. Циклины активируют CDK, что является сигналом к удвоению хромосом и делению клетки.
В процессе митотического деления каждая клетка получает полный двойной комплект хромосом. Если бы образование половых клеток шло таким же путем, то после оплодотворения (слияния женской и мужской гаметы) число хромосом всякий раз удваивалось бы. На самом деле этого не происходит. Каждому виду свойственно определенное число и определенный набор хромосом (кариотип). Это возможно лишь при условии, если при образовании клеток будет происходить редукция, т.е. уменьшение, числа хромосом вдвое.

Глава 4
Генный уровень организации наследственного материала. Химическая и структурная организация нуклеиновых кислот (ДНК и РНК). Репликация и репарация ДНК. Структура генов прокариот и эукариот. Генетический код и его свойства
Исследования, направленные на выяснение химической природы наследственного материала показали, что материальным субстратом генов являются нуклеиновые кислоты – ДНК и РНК. Нуклеиновые кислоты – это макромолекулы, т.е. отличаются большой молекулярной массой.
В структурной организации молекулы ДНК можно выделить 3 уровня:
Первичная структура – полинуклеотидная цепь
Вторичная структура – две комплементарные друг другу и антипараллельные полинуклеотидные цепи
Третичная структура – трехмерная спираль
Первичная структура днк
ДНК – это полимер, состоящий из мономеров – нуклеотидов. Каждый нуклеотид включает три основные компонента: азотистое основание (аденин, гуанин, цитозин, тимин, урацил), сахар (дезоксирибоза или рибоза), остаток фосфорной кислоты (рис. 4.1).
Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путём взаимодействия фосфата одного нуклеотида с гидроксилом д.р. так, что между ними устанавливается фосфодиэфирная связь. В результате образуется полинуклеотидная цепь. Важно отметить, что сборка полипептидной цепи осуществляется строго в одном направлении, а именно, путем присоединения фосфатной группы, расположенной в 5’-положении последующего нуклеотида к 3’гидроксильной группе предыдущего нуклеотида.
Вторичная структура днк
В 1953 г Уотсон и Крик представили модель 3-х мерной молекулы ДНК (рис. 4.2). За это открытие в последующем они были удостоены Нобелевской премии. Ученые показали, что особенностью II структурной организации ДНК является то, что в ее состав входят 2 полипептидные цепи, связанные между собой особым образом – путём образования водородных связей между азотистыми основаниями по принципу комплементарности: пурин связывается только с пиримидином, т.е. Аденин (А) может связываться только с Тимином (Т), а Гуанин (Г, англ. G) – только с Цитозином (Ц, англ. C) и наоборот. При этом между А и Т образуются 2 водородные
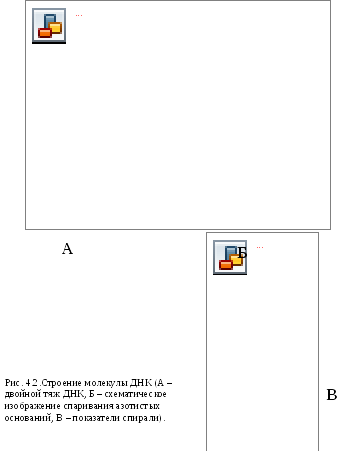
связи, а между Г и Ц – 3. Благодаря комплементарности соединения в цепь число пуринов в молекуле ДНК всегда равно числу пиримидинов, т.е.
A + Г = T + Ц – это правило Чаргаффа (1951 г.)
Хотя водородные связи между парами оснований относительно слабы, каждая молекула ДНК содержит приблизительно 3,3 млрд. пар, так что в физиологических условиях (Т0 , рН) цепи никогда не разрываются.
Сахаро-фосфатный остов находится по периферии молекулы ДНК, а пуриновые и пиримидиновые основания – внутри.
Другой важной особенностью молекулы ДНК является антипараллельность двух составляющих её цепей, т.е. 5’– конец одной цепи соединяется с 3’-концом другой.