

Тема 4. Основные уровни структурной организации живых организмов
История изучения основных уровней структурной организации живых организмов. Клеточная теория и ее современная модификация. Вирусоподобные: размеры, форма, строение, положение среди других живых систем. Бактериоподобные: размеры, форма, строение, положение среди других живых систем. Клеткоподобные или простейшие. Размеры, форма, строение, органы передвижения, положение среди других живых систем. Разноклеточные. Строение на клеточном и надклеточном уровнях. Ткани и органы. Макротаксономия и краткая характеристика основных таксонов – растений, грибов и животных.
История изучения основных уровней структурной организации живых организмов. Клеточная теория и ее современная модификация
Осознание того факта, что в природе имеются различные уровни структурной организации живых систем, пришло к ученым далеко не сразу. Накопление данных и их осмысление продолжалось несколько веков. Поэтому необходимо посвятить некоторое время истории этого вопроса.
Начнем с 1665 г., когда Роберт Гук изучал в микроскоп срезы пробки и, увидел, что она состоит из ячеек. Этим ячейкам он дал латинское название – cellula, что в переводе означает клетка. Он полагал, что клетки пусты, а живое вещество – это стенки. Сейчас нам понятно, что Р.Гук наблюдал всего лишь клеточные стенки. С тех пор длительное время внутреннему содержимому клеток не придавали большого значения. Антони ван Левенгук, М.Мальпиги, Н.Грю и другие исследователи под названиями пузырьков (vesicles), мешочков (utricili) или клеток в конце XVII века описали микроскопические структурные элементы, составляющие ткани у растений. Был обнаружен и мир микроскопических одноклеточных организмов, включая туманное описание в 1676 г. бактерий.
В XVIII веке интенсивный процесс изучения клеток и тканей был сильно заторможен Карлом Линнеем, т.к. великий биолог весьма скептически относился к микроскопическим исследованиям. Да и микроскопы тогда были не совершенны и при большом увеличении давали размытое изображение. В результате работы были свернуты, а понятие «клетка» забыто.
Усовершенствование микроскопа и разработка основ гистологических методик в начале XIX века возродили интерес к изучению клеток и тканей организмов. Французский
35
ученый Бриссо-Мирбэ снова стал использовать термин «клетка», он же в книге «Изложение теории организации растений» (1802 г.) сделал обобщение что, все растения образованы тканями, состоящими из клеток. Ж.Б.Ламарк в «Философии зоологии» распространил эту идею на все живые существа. В «Учебнике натурфилософии» (1809 г.) Л.Окен клетками считал как ячейки и пузырьки тканей растений и животных, так и микроскопических инфузорий. В 1824 г. вышла книга французского ученого Г.Дютроше «Анатомические и физиологические исследования о тонком строении животных и растений, а также о их подвижности», в которой он сделал выводы о том, что все ткани растений и животных состоят из клеток, которые являются индивидуумами, а формирование и рост тканей происходит за счет увеличения числа клеток.
В 1831 г. Роберт Браун описал в растительных клетках ядро (первым его наличие отметил в 1830 г. Я.Пуркинье (или Я.Пуркине)). Затем было описано деление клеток у ряда водорослей. Среди ученых начал обсуждаться вопрос о том, как образуются новые клетки.
Традиционно считается, что в 1838–1839 г.г. зоолог Т.Шванн и ботаник М.Шлейден объединили идеи разных ученых и сформулировали так называемую «клеточную теорию». Первоначально она имела следующий вид:
1.Все живые существа (а тогда различали только два царства – Растения и Животные) состоят из клеток.
2.Клетка – независимый индивид (организм), являющийся элементарной единицей живых существ.
3.Клетка – это пространство, ограниченное вегетативной мембраной (оболочкой) и содержащее слизистую жидкость и ядро. Главным компонентом клетки является оболочка, так как в ней совершаются основные метаболические процессы.
4.Все клетки возникают путем цитогенеза из цитобластемы, которая в тканях животных расположена главным образом между клетками (последний пункт – это представления М.Шлейдена).
Так как главные положения клеточной теории были достаточно строго сформулированы Дютроше до Шванна и, учитывая, что Шванн взял у Шлейдена лишь оказавшуюся неправильной гипотезу цитогенеза, выдающийся протистолог Л.Н.Серавин предложил, что исторически справедливо будет считать Дютроше как минимум соавтором клеточной теории. Кроме того, известно, что Шлейден был знаком с работами Дютроше. Есть доказательства того, что и Шванну они были знакомы, но он не ссылался на них.
36
Впрочем, клеточная теория Дютроше–Шванна была несовершенной. Так, вскоре было доказано, что внутреннее содержимое клетки является ее существенным компонентом. Моль дал ему название «протоплазма». Затем было установлено, что клетки размножаются путем деления, что позволило Р.Вирхову выдвинуть принцип: «всякая клетка — из клетки». Было также обнаружено, что все растительные клетки имеют тонкую полупроницаемую мембрану, которую нельзя смешивать с толстой неживой клеточной стенкой.
Впоследующие годы сформировался вариант классической клеточной теории. Ее положения таковы:
1.Все живые существа состоят из клеток (одной или многих).
2.Клетка – элементарная единица всего живого.
3.Любая клетка состоит из трех непременных компонентов: поверхностной мембраны, цитоплазмы и ядра, обладающего собственной оболочкой.
4.Каждая клетка происходит из клетки (путем митоза), а в более сильном варианте – все живое — из клетки.
Классическая теория была разработана на простейших, водорослях, высших растениях и животных. Существование бактерий и, главное, их отличие от других организмов тогда еще не было осознано. Лишь в конце XIX века заметили, что у бактерий нет ядра. Поэтому, когда Э.Геккель в 1866 г. выделил наряду с существовавшими царствами Plantae (Растения) и Animalia (Животные) третье царство – Protista, то в нем он выделил особый таксон безъядерных протистов – класс Monera (Монеры).
Для строгих приверженцев клеточной теории открытие доядерных форм живого казалось опровержением самой теории. Поэтому они сделали все возможное, чтобы доказать, что бактерии являются клетками. До сих пор во многих современных книгах и учебниках по-прежнему можно увидеть фразы «бактериальные клетки» или «прокариотные клетки». Это неверно по определению.
Вконце XIX века состоялось «открытие» вирусов. В 1892 г. русский ботаник Д.И.Ивановский впервые получил инфекционный экстракт из растений табака, пораженных мозаичной болезнью. Экстракт сохранял инфекционные свойства, даже будучи пропущен через фильтр, задерживающий бактерии. Голландец Бейеринк в 1898 г. придумал для обозначения неизвестного инфекционного начала таких жидкостей термин «вирус», от латинского слова virus – яд. Позднее установили, что очищенные пробы содержали нуклеопротеины, но сами частицы оставались неуловимыми. Лишь после изобретения электронного микроскопа удалось увидеть и исследовать сами вирусы, которые явно имели не
37
клеточное строение. Перед биологами встал вопрос: «Являются ли вирусы живыми?». Если живой считать структуру, обладающую генетическим материалом и способную себя воспроизводить, то тогда вирусы можно считать живыми. Разумеется, были предприняты попытки, считать вирусы клетками. Тем не менее, вероятно, большинство ученых сейчас вирусы относят как минимум к неклеточным формам жизни.
Первые три положения клеточной теории с открытием безъядерных организмов (бактерий) и внеклеточных форм жизни (вирусов) пришли в противоречие с новыми фактами. Что касается четвертого положения: все живое из клетки, то оно, как считает Л.Н.Серавин, не исторично. Ведь клетка не могла возникнуть сразу в законченном сложном виде.
Между тем в 1925 г. французский ученый Шаттон предложил новый подход к изучению структурной организации живых систем. Он разделил живые организмы на две группы: прокариот и эукариот (по-гречески karion – ядро; более правильно писать и произносить евкариоты, но эукариоты привычней). К первым он отнес организмы, не имеющие ядра, т.е. все бактерии. Ко вторым – организмы, имеющие ядра.
Предложенное Шаттоном деление живых систем в зависимости от уровня их структурной организации длительное время не находило поддержки, поскольку не согласовалось с клеточной теорией. Однако электронная микроскопия показала, что бактерии действительно лишены ядра. У них есть только нуклеоид с кольцевой молекулой ДНК. Догадка Э.Геккеля, что тело монеры еще не обособилось в протоплазму и ядро, оказалась правильной. Э.Геккель считал, что монера еще не есть клетка, и предложил называть их цитодами. В ходе дальнейших исследований обнаружились и другие отличия прокариот от эукариот. Например, у прокариот отсутствует митоз.
Как уже говорилось выше, клеточная теория разрабатывалась на простейших, водорослях и других организмах, которые все являются эукариотами. Поэтому Л.Н.Серавин считает, что клеточная теория применима только к эукариотам. Соответственно и термин «клетка» применим только к ним. Кроме того, научные термины должны быть однозначными. Если бактерию считать клеткой, тогда и такие клеточные органеллы как хлоропласты и митохондрии, имеющие собственную наружную мембрану, матрикс и нуклеоид, придется называть клетками.
Организм прокариот Л.Н.Серавин предлагает называть процитом (от лат. pro – до и cytus – клетка), так как бактерии по уровню структурной и функциональной организации еще не являются клетками. Процит состоит из трех основных компонентов: бактериомем-
38
браны, бактериоплазмы и нуклеоида. Структурные компоненты бактерий не являются прямыми гомологами трех основных компонентов настоящих клеток.
Что касается структурной единицы эукариот, то здесь сначала необходимо рассмотреть ряд моментов. В соответствии с клеточной теорией организмы, относимые к Protozoa (Простейшие), считались одноклеточными, а Metazoa, соответственно, многоклеточными. Но новые данные говорят о том, что среди Простейших есть как одноклеточные (моно- и полиэнергидные), так и колониальные и многоклеточные организмы (но находящиеся на дотканевом уровне организации). Энергида — структурная единица клетки, представляющая собой ядро с прилегающим участком цитоплазмы и органеллами, но без клеточной оболочки. Среди полиэнергидных (многоядерных простейших) различают симпласты, не меняющие форму тела и плазмодии, способные амебоидно менять форму своего тела. Все одноклеточные простейшие устроены сложнее, чем любая клетка у Metazoa. Поэтому, например, весьма сложно устроенные организмы инфузорий обозначают термином не клетка, а сомателла. В тоже время многоклеточные организмы (как животные, так и растения) могут состоять не только из сравнительно просто устроенных, пусть специализированных клеток, но и из синцитиев. Следовательно, многоклеточные организмы не всегда состоят только из обычных клеток и правильнее их называть гетероклеточными. Для обозначения всех структурных единиц эукариот Л.Н.Серавин предложил термин «цитоиды». Соответственно настоящая клетка это эуцит. А сомателлы, синцитии, симпласты и плазмодии – это сложные цитоиды.
Утверждение, что клетки обязательно происходят от клетки, не подтверждается. Так сложные цитоиды могут распадаться на эуциты без митотического деления ядер. Однако все эукариотические организмы на какой-то стадии жизненного цикла проходят одноклеточную стадию, что, вероятно, свидетельствует об их происхождении от одноклеточного предка. Поэтому назрела необходимость в изменении классической клеточной теории в соответствии с новыми фактами. И в 1991 г. Л.Н.Серавин предложил цитоидную теорию организации эукариот, являющуюся современной модификацией клеточной теории. Суть
еезаключается в следующих положениях:
1.Все эукариотические организмы состоят из цитоидов. Филогенетически первичной, наиболее простой и самой распространенной формой цитоида является клетка (эуцит).
2.Главными и непременными структурными компонентами клетки являются плазмалемма, цитоплазма и имеющее оболочку ядро. Сложные цитоиды (плазмодии, сомателлы и симпласты) имеют эти же компоненты и возникают в результате слияния нескольких
39
или многих клеток, на основе многократных, но до конца не доведенных делений клетки, а также благодаря сильной полимеризации органелл клетки и их комбинаций.
3. Каждый цитоид возникает из цитоида, чаще всего (филогенетически и онтогенетически) из клетки.
Следовательно, в природе все известные организмы распределяются по двум кластерам: Prokaryota и Eukaryota. Им соответствуют четыре уровня структурной организации. В кластере Prokaryota это вириоидный (Virioidata) и бактериоидный (Bacterioidata) уровни. В кластере Eukaryota – клеточный (или цитоидный Cellulata = Cytoidata) и гетероцитоидный уровни (Heterocytoidata). В процессе эволюции новый уровень структурной организации может достигаться независимо разными систематическими группами организмов.
Вирусоподобные: размеры, форма, строение, положение среди других живых систем
Вирусы – неклеточные формы жизни. Это наиболее просто устроенные и эффективные внутриклеточные паразиты, являющиеся возбудителями болезней всех групп организмов: от бактерий до гетероклеточных растений и животных. Можно сказать, что вирусы — это внутриклеточные паразиты на генетическом уровне.
История обнаружения вирусов была изложена выше. Поэтому перейдем к описанию строения вирусов.
Вирусы существуют в двух формах: покоящейся или внеклеточной (называются вирусными частицами или вирионами) и репродуцирующейся или внутриклеточной (комплекс вирус–клетка).
В свободном состоянии (вне клетки) простые вирусы (рис. 1) состоят из молекулы нуклеиновой кислоты и белковой защитной оболочки – капсида (от лат. capsa – вместилище), часто образованной из идентичных повторяющихся субъединиц – капсомеров. Некоторые вирусы способны кристаллизоваться. Их форма – палочковидная, нитевидная или сферическая. Сложные вирусы – герпес, грипп и др. (рис. 1, 2) могут иметь дополнительную белковую или липопротеиновую оболочку (возникает из плазматической мембраны клетки-хозяина), иногда содержащую еще и углеводы.
Размер вирионов 15–350 нм (иногда до 2000 нм), т.е. в среднем они раз в 50 меньше бактерий.
40
Рис. 1. Структура вируса табачной мозаики (А) и форма вирусных частиц герпеса (Б) и гриппа (В) (из Мамонтов, Захаров, 1999): 1– капсид, 2 – РНК.
Рис. 2. Строение вириона ВИЧ по Галло, Монтанье (1988) (из Кусакин, Дроздов, 1997): ЛДМ – липидная двухслойная мембрана, Р – ревертаза.
41
По-видимому, в вирусах присутствует всегда один тип нуклеиновой кислоты – либо ДНК, либо РНК. Молекулы нуклеиновых кислот могут быть одноили двухцепочными, линейной или кольцевой структуры, иногда в виде набора фрагментов, заключенных в разные полые структуры, окруженные белковой оболочкой. Размеры молекул варьируют от четырех генов у РНК-содержащего бактериального вируса Qβ и до примерно 250 генов у вируса оспы.
Описано более 1000 ДНКили РНК-содержащих вирусов, поражающих теплокровных животных (из них половина патогенна и для человека), и около 600 вирусов, поражающих растения (все РНК-содержащие). Известны и вирусы бактерий – бактериофаги (рис. 3) или просто фаги (от греч. «фагео» – поедать). Они изучены лучше других вирусов.
Итак, вирусы – облигатные паразиты. Самостоятельно размножаться они не способны. Все активные процессы вирусов протекают в клетках-хозяевах (рис. 4). В клетку генетический материал вируса попадает в процессе, напоминающем инъекцию. Белковая оболочка вируса остается на внешней поверхности клетки (есть мнение, что процесс проникновения может быть различным). По характеру репродукции различают лизогенезирующие и лизирующие бактериофаги. После попадания нуклеиновой кислоты лизирующего вируса в бактерию синтезируются иРНК, кодирующие ферменты, которые настраивают работу аппарата синтеза пораженного процита на производство молекул вирусной нуклеиновой кислоты и оболочки вируса. Далее, в результате самосборки формируются дочерние вирусные частицы. Бактерия гибнет, лизируется (лопается под действием фермента лизоцима) и высвобождается 200–1000 новых фагов. В случае лизогенезирующего вируса (например, фаги λ или Τ4) нуклеиновая кислота не сразу репродуцируется, а сначала встраивается в определенный участок ДНК бактерии, оставаясь профагом или, по-другому, провирусом (т.е. не активным вирусом). Профаг ведет себя как часть бактериального генетического материала, реплицируясь в течение нескольких поколений в каждом цикле деления. Такие фаги известны под названием умеренных фагов, а бактерии, в которых они затаились, называют лизогенными. В определенных условиях профаг выходит из ДНК бактерии и начинает синтез вирусных частиц, вступая в лизирующую фазу жизненного цикла. К встроенным вирусам относятся многие онкогенные вирусы растений и животных.
Известны вирусы-сателлиты. Их одноцепочечная молекула ДНК способна реплицироваться лишь в присутствии другого вируса. Пример: РНК-вый S-вирус, реплицируется лишь в присутствии вируса гепатита B.
42
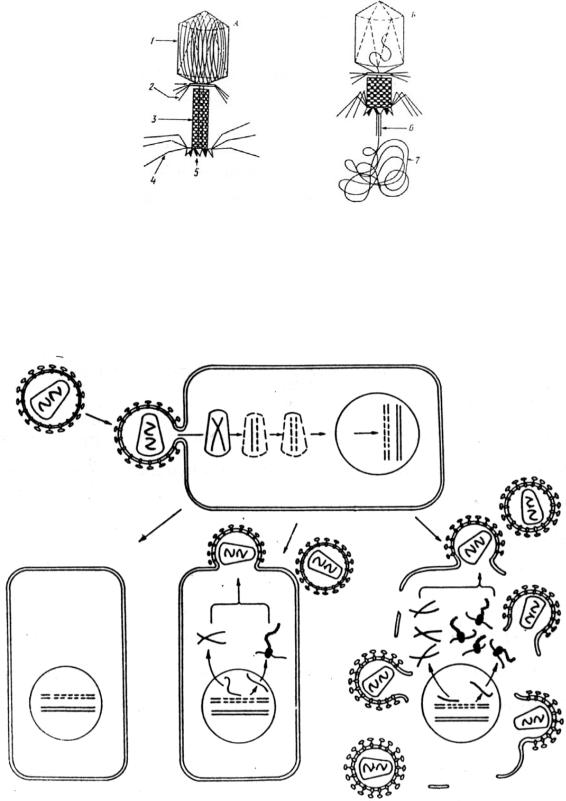
Рис. 3. Модель бактериофага Т-2 (из Мамонтов, Захаров, 1999). А – фаг в момент прикрепления к поверхности бактерии; Б – фаг во время введения ДНК в процит: 1 – головка, содержащая ДНК, 2 – воротничок, 3 – чехол ножки, 4 – хвостовые нити, 5 – базальная пластинка, 6 – стилет, 7 – вводимая ДНК.
Рис. 4. Жизненный цикл ВИЧ (из Кусакин, Дроздов, 1998).
Инфекция начинается со слияния оболочки вируса с плазмалеммой и проникновения сердцевины вируса в цитоплазму. Обратная транскриптаза синтезирует одноцепочечную ДНК вируса (пунктир), комплементарную вирусной РНК. Вирусная ДНК (провирус) проникает в ядро клетки и встраивается в хромосомную ДНК. При активации провируса, на нем транскрибируется мРНК, затем идет синтез РНК-копий и вирусных белков, с последующим формированием новых вирусных частиц.
43
К вирусам относят и вирусоиды – небольшие кольцевые молекулы РНК, заключенные в белковый капсид, и способные реплицироваться только в присутствии вирусовпомощников.
Вироиды – РНК-вые агенты, представленные молекулой РНК длиной около 360 нуклеотидов, в которой нельзя закодировать даже один белок средних размеров. Вызывают болезни растений.
Особую группу составляют агенты белковой природы, вызывающие медленные инфекции (губчатые энцефалопатии) с длительным инкубационным периодом. Разрушают нервные клетки, вызывают паралич. Предполагают, что такие белки служат активаторами своих же генов, заставляя их вырабатывать все новые порции продукта, накопление которого приводит к болезни. Пример: возбудитель скрепи — неврологической болезни овец или заболевание мозга «куру» среди членов племени «охотников за головами» на Новой Гвинее (инкубационный период 20 и более лет).
Имеется несколько гипотез происхождения вирусов:
1.Согласно гипотезе о первичном происхождении вирусных систем, современные вирусы — отражение доклеточного этапа эволюции.
2.Другая гипотеза отстаивает идею регрессивной эволюции вирусов из бактерий. Сторонники данной гипотезы считают вирусы конечными пунктами дегенерации клеток. Например, по схеме: бактерии → риккетсии → хламидозоа → вирус. Или предполагают, что вирусы развились из органоидов клеток: митохондрий, хлоропластов, эписом.
3.Наибольшее число сторонников имеет гипотеза о происхождении вирусов из клеточных генетических структур. В пользу этой гипотезы говорит возможность синтеза инфекционных вирусов из простых предшественников – матричной нуклеиновой кислоты и ферментов. Возможно, что вирусы возникали многократно в ходе эволюции органического мира и что они имеют полифилетическое происхождение. Тогда естественную систематику вирусов создать будет весьма сложно.
Обычно при классификации вирусов учитывают тип нуклеиновой кислоты, природу хозяев или их биохимические, морфологические и физиологические особенности. Сейчас все вирусы объединяют в царство Vira или Virae (некоторые систематики считают, что необходимо выделить несколько царств вирусов, тогда отдельные царства объединяют в империю Noncellulata). В царство Vira входят два типа: Тип Ivanovskia (РНК-содержащие вирусы) и Тип Jenneria (ДНК-содержащие вирусы). К настоящему времени выделено около 60 семейств вирусов животных, 26 групп вирусов растений.
44
Для названий семейств вирусов используют окончания -viridae (например, Poxviridae). Для родов окончание -virus (например, Enterovirus). Для видов биноминальные названия не прижились и названия вирусов, как правило, тривиальные. Например: вирус табачной мозаики, бактериофаг Х–174, ВИЧ, вирус полиомиелита.
Бактериоподобные: размеры, форма, строение, положение среди других живых систем
Кбактериоподобным относят следующие организмы: настоящие бактерии, синезеленые водоросли (цианобактерии), микоплазмы, актиномицеты, риккетсии и др. организмы, находящиеся на прокариотном уровне организации.
Ксобственно бактериям (от греч. bakterion – палочка), традиционно относят прокариотные микроорганизмы, состоящие из одного процита, или объединенные в организованные группы, которые представляют собой кокки или палочки, неподвижные или со жгутиками, противопоставляя их морфологически более сложным прокариотам – актиномицетам, цианобактериям, миксобактериям. Размер бактерий в среднем около одного микрометра (варьируют от 0,1 до 10 мкм), что на порядок меньше клеток эукариот (объем меньше в 1000–10000 раз). Диаметр кокков не превышает 0,5 мкм. Среди микоплазм есть организмы размером менее 0,125 мкм, т.е. они меньше некоторых вирусов. По форме различают следующие бактерии: сферические – кокки; палочковидные – бациллы; спиралевидные – спириллы (спирально извитые палочки с полярным жгутиком) и спирохеты (закручены винтообразно, имеют фибриллы); короткие, изогнутые в виде запятой палочки – вибрионы.
Главная особенность организма прокариот (рис. 5) – отсутствие ядра. Молекула ДНК (5·106 пар нуклеотидов) не окружена мембраной, а в виде кольцевой хромосомы (генофор – состоит из ДНК, РНК и белков) расположена в нуклеарной области бактериоплазмы (вместе они называются нуклеоидом) и закреплена на бактериомембране. ДНК прокариот связана с очень небольшим количеством белка. В среднем она содержит несколько тысяч генов. Вне нуклеоида имеются плазмиды – небольшие кольцевые молекулы ДНК, число которых варьирует, а утрата не сказывается на репродукции клетки. Плазмиды могут придавать своим хозяевам особые свойства, например, устойчивость к антибиотикам. Внутри процит не разделен мембранами на отделы или отсеки (не компартментализован). Из внутренних мембран может быть мезосома, функция которой не вполне ясна.
45
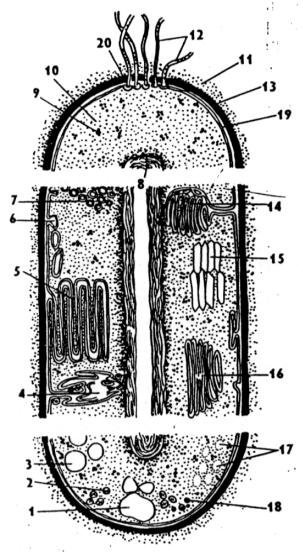
Рис. 5. Внутреннее строение бактерии (из Яковлев, Аверьянов, 1996):
1– гранулы поли-β-оксимасляной кислоты, 2 – жировые капли, 3 – включения серы, 4 – трубчатые тилакоиды, 5 – пластинчатые тилакоиды, 6 – выросты бактериомембраны, 7– хроматофоры, 8 – нуклеоид, 9 – рибосомы 10 – бактериоплазма, 11 – слизистая капсула, 12 – жгутики, 13 – бактериальная оболочка, 14 – мезосома, 15 – газовые вакуоли, 16 – ламеллярные структуры, 17 – гранулы полисахаридов, 18 – гранулы полифосфатов, 19 – бактериомембрана, 20 – базальное тельце.
46
Возможно, что она принимает участие в синтезе материала для бактериомембраны после деления процита, в дупликации ДНК перед делением бактерии или в клеточном дыхании. Есть мнение, что мезосома вообще отсутствует в нормальных бактериях, т.е. является артефактом, появляющимся в экстремальных условиях. Фотосинтезирующие бактерии имеют мембраны, которые содержат молекулы, необходимые для фотосинтеза, но они не заключены в хлоропласты. По всей бактериоплазме располагается до 10000 более мелких, чем у эукариот рибосом (70S-рибосомы, где S (сведберг) – единица, характеризующая скорость седиментации в центрифуге; чем больше число S, тем выше скорость седиментации) и иногда видны включения запасных питательных веществ. Митохондрий, аппарата Гольджи, вакуолей, лизосом, центриолей, пластид нет. Отсутствует движение бактериоплазмы внутри процита. Нет фагоцитоза и пиноцитоза. От внешней среды содержимое процита отделяет бактериомембрана. Снаружи находится защитная оболочка – бактериальная стенка. Выделяют четыре типа строения стенки тела прокариоты:
1.Фирмикутный тип – характерен для грамположительных бактерий. Клеточная стенка на 30–70 % (сухой массы) состоит из особого гетерополимера – пептидогликанмуреина (встречается только у прокариот). С наружной стороны с муреином связаны тейхоевые кислоты. Такие комплексы образуют несколько слоев (до 40), сшитых между собой
вгигантскую макромолекулу. Аминокислотный состав муреина видоспецифичен.
2.Грациликутный тип – только у грамотрицательных бактерий. Один–два слоя муреинового каркаса (всего 10 % от массы стенки), покрытого снаружи сложным комплексом, содержащим белки, фосфолипиды, липопротеиды, липосахариды. Комплекс имеет двухслойную структуру, напоминающую структуру клеточных мембран. Поэтому ее называют наружной бактериальной мембраной. Она служит барьером проницаемости для различных веществ. В мембране могут содержаться порины – крупные трансмембранные белки, имеющие гидрофильные поры для пропуска гидрофильных низкомолекулярных биомолекул. Между муреиновым слоем и бактериоплазматической мембраной есть периплазматическое пространство. В нем находятся ферменты и связующие белки, обеспечивающие перенос макромолекул в бактериоплазму.
3.Мендозикутный тип – характерен для архебактерий. Главная особенность: отсутствие мурамовой кислоты и муреина (встречается псевдомуреин), с чем связана устойчивость к действию антибиотиков, ингибирующих синтез муреина (пенициллин, цефалоспорин). Архебактерии отличаются от других организмов еще и однослойным строением ли-
47
пидных бактериоплазматических мембран, в которых нет жирных кислот (есть простые эфиры глицерина и нейтральные изопреноидные углеводороды).
4. Тенерикутный тип – характерен для микоплазм, в том числе и для термоплазмы, у которой бактериальная стенка как таковая отсутствует, а организм окружен только бактериоплазматической мембраной.
Бактериальная стенка может быть покрыта капсулой или слизистым слоем – это слизистые или клейкие выделения некоторых бактерий, которые служат дополнительной защитой (например, некапсулированные штаммы пневмококков легко атакуются и уничтожаются фагоцитами) или формируют колонии из отдельных процитов. Капсула – относительно толстое и компактное образование, слизистый слой – намного рыхлее.
Многие бактерии подвижны (за одну секунду они могут преодолевать расстояние, равное 20 диаметрам процита). Эта подвижность обусловлена выделением слизи или наличием одного или нескольких жгутиков. По строению жгутики бактерий устроены гораздо проще, чем у эукариот. По структуре они напоминают одну из микротрубочек эукариотического жгутика. Состоят из сферических субъединиц белка флагеллина (похож на мышечный актин). Субъединицы расположены по спирали и образуют полый цилиндр диаметром около 10–20 нм. Основание жгутика, по-видимому, вращается так, что жгутик как бы ввинчивается в среду, продвигая бактерию вперед. Это, наверное, единственная в живой природе структура, где используется принцип колеса. Для субъединиц флагеллина известно свойство спонтанно собираться в растворе в спиральные нити (самосборка!). Подвижные бактерии в ответ на определенные раздражения могут передвигаться. Например, подвижные фотосинтезирующие бактерии плывут к свету, аэробные бактерии обладают положительным аэротаксисом.
На стенках тела некоторых грамотрицательных бактерий есть тонкие нитевидные белковые (белок пилин) выступы – фимбрии (от лат. fimbriae –бахрома) или пили (F- ворсинки, половые волоски, копуляционные фимбрии). Их число варьирует от одной до двухсот. Служат для прикрепления бактерий друг к другу или к субстрату. F-пили кодируются специальной плазмидой и связаны с «половым» размножением бактерий.
Отношение поверхность/объем у бактерий очень велико. Это дает возможность быстро поглощать питательные вещества за счет диффузии и активного транспорта. В благоприятных условиях бактерии растут очень быстро. Достигнув определенных размеров, бактерии приступают к бесполому размножению – бинарному делению (митоз и мейоз у прокариот отсутствуют). Перед делением ДНК реплицируется, образуя две идентичные
48
молекулы, при этом мезосомы удерживают геном в определенном положении. Затем бактериомембрана врастает между этими двумя молекулами ДНК. И в каждой дочерней бактерии оказывается по одной молекуле ДНК. У самых быстрорастущих бактерий деление может происходить каждые 20 минут. Интервал между делениями называется временем генерации.
У бактерий есть и примитивный «половой» процесс (меромиксис). Конечно, бактерии не образуют гамет, с последующим оплодотворением. Но в процессе генетической рекомбинации у них может происходить обмен генетическим материалом. Часть ДНК (редко вся ДНК) бактерии-донора переносится в бактерию-реципиент, ДНК которой генетически отличается от ДНК донора. Перенесенная ДНК замещает часть ДНК реципиента. Образуется рекомбинантная ДНК. Она содержит гены обеих бактерий. Этот процесс очень важен для выживания и эволюции бактерий, так как повышает наследственную изменчивость.
Известны три способа получения рекомбинантов:
1.Трансформация (от лат. transformatio – преобразование, превращение) – процесс, при котором из бактерии-донора выходит небольшой фрагмент ДНК, поглощается реципиентом и включается в состав его ДНК, замещая похожий, хотя и не обязательно идентичный фрагмент. Открыл трансформацию в 1928 г. Ф.Гриффит в экспериментах с пневмококками. К ней способны лишь немногие бактерии, называемые «компетентными».
2.Конъюгация (от лат. conjugatio – соединение) – это однонаправленный перенос ДНК между бактериями, непосредственно контактирующими друг с другом посредством F-фимбрий. Может обмениваться значительная часть донорской ДНК. Процесс открыт в 1946 г. в эксперименте с Escherichia coli. Донорская способность бактерии определяется генами, находящимися в небольшой кольцевой молекуле ДНК. Ее называют половым фактором или F-фактором (F – первая буква от англ. fertility – плодовитость). F-фактор – это своеобразная плазмида, кодирующая белок F-пилей. При конъюгации одна из цепей двухцепочечной ДНК F-фактора проникает через половую фимбрию из бактерии-донора в бак- терию-реципиент. И у донора и у реципиента молекулы ДНК реплицируются. Постепенно вся популяция бактерий становится носителем F-фактора. Бактерии-реципиенты могут иногда спонтанно утрачивать F-фактор. Очень редко (в одном случае из ста тысяч) F- фактор встраивается в молекулу основной ДНК бактерии. Тогда при конъюгации переносится не только F-фактор, но и остальная ДНК. Процесс занимает около 90 минут. Если бактерии расходятся раньше, то передается лишь часть ДНК. Штаммы, которые постоянно
49
передают всю или большую часть своей ДНК другим бактериям, называют Hfr-штаммами
(от англ. H = hidh – высокая, f = frequency – частота, r = recombination – рекомбинация).
3. Трансдукция – процесс, при котором небольшой двухцепочечный фрагмент ДНК попадает из бактерии-донора в бактерию-реципиент вместе с бактериофагом. ДНК некоторых вирусов может встраиваться в ДНК бактерий и передаваться из поколения в поколение. Иногда вирусная ДНК активируется и начинает кодировать образование новых фагов. ДНК бактерии-хозяина разрывается, и фрагменты иногда захватываются внутрь вирусных частиц (порой вытесняя ДНК самого вируса). Такие вирусы или трансдуцирующие частицы могут переносить ДНК в другие бактерии.
Физиология бактерий по разнообразию превосходит физиологию всех остальных форм жизни. Для получения энергии они используют различные органические и неорганические соединения (хемотрофы) или солнечный свет (фототрофы). Среди внутриклеточных паразитов имеются так называемые энергетические паразиты, использующие энергодающие реакции хозяина. Бактерии в состоянии разлагать практически все природные соединения, как в окислительных реакциях, так и анаэробно. Бактерии участвуют в круговороте всех биологически важных веществ в биосфере. Только они осуществляют ключевые реакции круговорота веществ: нитрификацию, денитрификацию, азотфиксацию, окисление и восстановление соединений серы. Ежесуточно бактерии перерабатывают около 5 млн. т сульфатов, производя около 1,5 млн. т сероводорода. Метанобразующие бактерии дают ежегодно 5–10·108 т метана. В процессах деструкции роль бактерий — определяющая.
Большинство бактерий космополиты: встречаются по всей планете и в разных экологических условиях. В одном грамме почвы может быть до 100–2500 млн. бактерий, а в одном миллилитре парного молока – до трех миллиардов. Бактерии способны существовать в крайне неблагоприятных для других организмов условиях. Например, оптимальные температуры для галофильных и термофильных бактерий лежат в диапазоне 85–90°C (выдерживают до 105°C). Чтобы пережить экстремально неблагоприятные условия, они способны образовывать внутри процита эндоспоры (споры такого типа у эукариот неизвестны). Прокариоты — самые древние организмы. Именно они миллиарды лет назад сформировали биосферу, сходную с современной. Особую роль в этом сыграли фотосинтезирующие цианобактерии (возможно, что процесс фотосинтеза возник 3,3 млрд. лет назад).
Для построения систем микроорганизмов используют два подхода. Первый заключается в составлении каталога форм, удобного для практического распознавания организмов.
50
Следовательно, это искусственная система, в которой могут использоваться различные признаки. Так, в 1884 г. К.Грам разработал метод окраски фиксированных бактерий в темно-фиолетовый цвет основными красителями (метиленовый синий, кристаллический фиолетовый и др.), с последующей обработкой йодом. После обработки спиртом одни бактерии обесцвечиваются – их называют грамотрицательными, другие сохраняют окраску, их называют грамположительными. Сейчас ясно, что такие свойства определяются различиями в строении стенок тел бактерий. Естественная или филогенетическая система прокариот разработана мало. Во-первых, это связано с недостатком палеонтологических данных. Во-вторых, по мнению многих исследователей, для бактерий в принципе невозможно построить филогенетическую систему, поскольку у них распространен обмен генами даже между представителями разных таксонов (например, при посредстве вирусов). Эта теория сетчатой эволюции лежит в основе фенотипической систематики бактерий, когда для построения системы используется комбинативный анализ сочетания многих фенотипических признаков. Другие ученые считают, что процессы переноса генов между представителями разных групп бактерий наблюдаются в основном в лабораториях, и переносить полученные данные на природные поселения пока рано.
В большинстве систем выделяют следующие таксоны (в ранге царства или подцарства) прокариот: Архебактерии (описано около 50), Настоящие бактерии или просто Бактерии (сюда относят большинство известных видов), Оксифотобактерии (включают цианобактерий, они же синезеленые водоросли (не менее 2000 видов) и хлороксибактерии).
Настольной книгой микробиологов мира в настоящее время является каталог, который называется «Определитель бактерий Берги». Названия бактерий даются в нем по бинарной номенклатуре.
Клеткоподобные или простейшие. Размеры, форма, строение, органы передвижения, положение среди других живых систем
В 1866 г. Э.Геккель выделил царство Protista (Протисты), включив в него бактерий, простейших, одноклеточные водоросли, низшие грибы и губки, причем безъядерные протисты составили отдельный класс Monera. Позднее, в процессе уточнения объема этого царства Э.Геккель распределил протистов между царствами животных и растений. Идею протистов возродил в 1956 г. Коупланд, включив в это царство только эукариотические организмы, в том числе настоящие грибы. В современном объеме, как одно из четырех
51
царств эукариот, протисты были введены в 1969 г. Р.Уиттекером. По Р.Уиттекеру протисты являются переходной в эволюционном плане группой эукариот от прокариотических организмов к другим эукариотам с многоклеточной и мицеальной организацией. Среди протистов можно найти все три типа питания, известные у эукариот: гетеротрофы (животные), автотрофы (растения) и сапрофиты (грибы). Известно немало организмов со смешанным типом питания (в зависимости от внешних условий) – миксотрофов. Они объединяют в себе признаки характерные и для растений и для животных. Споры по поводу, правомочности выделения отдельного царства Протистов и по его составу не утихают. В настоящее время насчитывают, по разным оценкам, от 129 до 200 тыс. видов этих организмов. Вероятно, что описана лишь меньшая часть действительно существующего в биосфере многообразия протистов. Они живут во всех возможных для жизни средах обитания. Среди них много паразитов. Например, есть основания полагать, что почти каждый вид насекомых и других членистоногих имеет свой специфический вид таких паразитов, как микроспоридии (т.е. число видов микроспоридий возможно превышает миллион!).
Гетеротрофные протисты составляют Подцарство Protozoa – Простейшие (в некоторых системах вы найдете их как подцарство в царстве Animalia). Изучает простейших наука протозоология. Весьма вероятно, что в систематическом отношении это сборная группа полифилетического происхождения, которая через окрашенных жгутиконосцев тесно связана с автотрофными протистами (водоросли), а через микроспоридий с грибами (рис. 6). Систематика простейших разработана недостаточно, поэтому назовем лишь некоторые таксоны: Тип Dinophyta – Панцирные жгутиконосцы (около 300 видов), Типы Euglenophyta и Chlorophyta – Зеленые эвглены и Хлорофитовые (ботаники традиционно относят их к водорослям, так как сюда входят и фотосинтезирующие организмы), Тип Sporozoae – Споровики (более 4000 видов облигатных паразитов беспозвоночных и позвоночных животных), Тип Ciliata – Инфузории (наряду с амебами, наиболее известная группа простейших, насчитывающая от 5500 до 7500 видов), Надтип Rhizopoda – Корненожки. Количество известных видов простейших по разным источникам составляет 50000 – 70000.
Раньше простейших называли одноклеточными организмами. По современным данным среди них есть как одноклеточные, так и колониальные (Род Volvox) и многоклеточные формы (Класс Myxosporidia, Тип Labirinthomorpha). Но все они находятся на дотканевом уровне организации. Т.е. большинство дифференцировок, приводящих к возникновению определенного плана организации, происходят внутри клеток. Гетероклеточность у них никогда не возникает (тканей нет!).
52
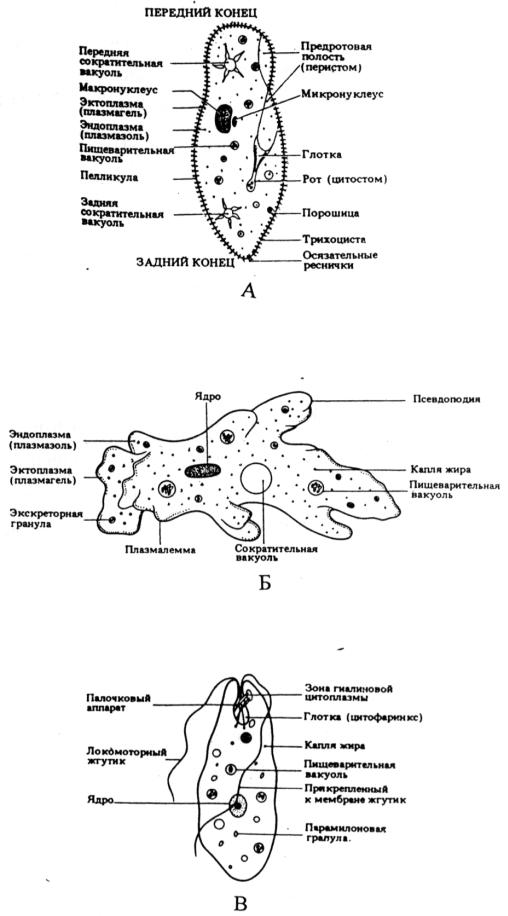
Рис. 6. Представители подцарства Protozoa (из Грин, Стаут, Тейлор, 1996):
А – Инфузория туфелька, Б – Амеба протей, В – Peranema trichophorum.
53
В качестве элементарной единицы организации простейших выделяют, так называемую, энергиду. Энергида – это участок клетки, включающий ядро, окружающую его цитоплазму с органеллами, но без пограничного слоя. Пограничный слой – это слой цитоплазмы, прилегающий к плазмалемме. Если клетка имеет одно ядро, то такое простейшее будет моноэнергидным. Если ядер много – то полиэнергидным (например, многоядерные амебы). Радиолярий, у которых ядро имеет полиплоидный набор хромосом, относят к скрытно полиэнергидным простейшим. Многоядерное тело, способное к амебоидному движению называют плазмодием, не способное – симпластом. Сложно устроенный организм инфузорий называют сомателлой (отметим, что вследствие того, что даже одноклеточные простейшие являются организмами и индивидуумами, то их клетки устроены сложнее клеток гетероклеточных организмов).
Средние размеры простейших менее одного миллиметра. Большинство видов имеют размер 5–150 мкм. Самый мелкий известный представитель подцарства – Babesia boa, внутриклеточный паразит крупного рогатого скота. Паразитический жгутиконосец Leischmania donovani чуть крупнее – 14 мкм. Размеры дизентерийной амебы Entamoeba histolytica 15–30 мкм. Инфузория туфелька Paramecium caudatum может достигать в длину
150–280 мкм, а амеба протей Amoeba proteus до 600 мкм. Длина инфузории Spirostomum ambiguum 3 мм. Некоторые колониальные радиолярии из рода Collsomum в длину бывают 10 – 20 см. Известны раковины вымерших радиолярий диаметром 16 – 18 см.
Микроскопические размеры большинства простейших определяют ограничения в освоении ими различных сред и биотопов. Следствием малых размеров является обширная относительная поверхность тела. С одной стороны это облегчает процессы обмена веществ через поверхность тела. С другой стороны простейшие подвержены быстрому высыханию. Поэтому простейшие обитают в воде или в среде с большим содержанием влаги. Противостоять неблагоприятным условиям (пересыханию мест их обитания) они могут благодаря способности образовывать цисты – покоящиеся стадии, снабженные защитной оболочкой. Цисты могут перемещаться ветром или другими организмами на большие расстояния, благодаря чему многие простейшие являются организмами-космополитами. Форма тела и окраска простейших чрезвычайно разнообразны. Скорость движения амеб находится в пределах 0,2-4,0 мкм в секунду, жгутиконосцев 30-325 мкм в секунду. Быстрее всех могут передвигаться инфузории – 200-3740 мкм в секунду. Вода для простейших является довольно вязкой средой, что затрудняет целенаправленное движение из-за крутящих гидродинамических моментов. Простейшие решили эту проблему путем стабилизации положения тела
54
вращением вокруг продольной оси. В толще воды они движутся по спиральным траекториям.
В зависимости от способов передвижения выделяют два основных типа организации клеток всех протистов: жгутиконосный (монадный) тип и амебоидный (саркодовый) тип.
Порой обе формы могут представлять собой разные стадии жизненного цикла.
Для монадного типа характерно наличие жгутиков (жгутик по латыни flagella) или ресничек. Эти системы используются для передвижения, временного или постоянного прикрепления к субстрату или для создания потоков воды, несущих пищу. У большинства современных жгутиконосцев два жгутика. Вероятно, предковые формы тоже имели два жгутика. Представления о системе Protista во многом базируются на строении жгутиков.
Структурной единицей локомоторного аппарата жгутиконосцев является мастигонт (2–4 кинетосомы со жгутиками). На этой основе выделяют четыре морфотипа жгутиконосцев. Изоконты (например, подвижные клетки зеленых водорослей) имеют от двух до восьми жгутиков равной длины, направленных в одну сторону, с одинаковыми способами биения. Анизоконты (бесцветные жгутиконосцы) несут два жгутика неравной длины, направленные в одну сторону, но отличающиеся по способу биения. Гетероконты (подвижные клетки водорослей, бесцветных жгутиконосцев и зооспоровых грибов) имеют два жгутика неравной длины, с разными способами биения. У стефаноконтов (многожгутиковые гаметы, зооспоры некоторых зеленых водорослей) на переднем конце клетки венчик жгутиков. Многие одножгутиковые формы рассматриваются, как утратившие вторично жгутик особи.
Жгутиковый аппарат состоит из свободной части (жгутик или ундулиподия), кинетосомы (центриоль) и корешковой системы. Тонкое строение жгутиков всех эукариот однотипно в главных чертах. Ундулиподия благодаря ритмичным сокращениям вызывает движение клетки или ток воды возле нее. Она представляет собой трубчатый вырост клеточной поверхности, окруженный мембраной, являющейся продолжением мембраны, покрывающей всю клетку. Ундулиподия содержит пучок белковых фибрилл (аксонему). Аксонема или аксиальная нить состоит из двух центральных микротрубочек, окруженных кольцом из девяти пар микротрубочек (дублетов). Микротрубочки образованы плотно спаянными друг с другом субфибриллами A и B. От A-трубочек в сторону соседнего дублета отходят пары «ручек» из белка динеина, а к центральным микротрубочкам тонкие филаменты (от позднелат. filamentum – нитевидное образование, нить), называемые спицами. Поверхность ундулиподии у свободноживущих форм опушена волосками (мастигонемы)
55
различных типов. Кинетосома похожа на цилиндр, стенки которого образуют девять триплетов микротрубочек. В проксимальной части цилиндра обычно расположена колесовидная структура: центральная ось связана с триплетами с помощью «спиц». Чаще всего имеются две кинетосомы, расположенные под прямым углом друг к другу и связанные между собой фибриллярными мостиками. Корешковая система – это совокупность микротрубочек или микрофиламентов, прикрепляющихся к кинетосомам или к ядру (кариомастигонт). Отростки корешков формируют под покровами (пелликулой) цитоскелет. Пелликула жгутиконосцев относительно плотная, укреплена различными структурами, что связывают с необходимостью создания опоры для локомоторного аппарата. Плотная пелликула привела к появлению, особенно у хищных видов, к появлению специализированных участков покровов, где происходит захват пищи (глотка и клеточный рот – цитостом) и выделение переваренных остатков (цитопрокт).
Для многих жгутиконосцев характерно наличие глазка или стигмы. Эта органелла, по-видимому, играет либо роль ширмы, затеняющей фоторецептор, либо рефлектора, усиливающего свет, либо экрана, защищающего фоторецептор от отраженных сигналов изнутри клетки.
Амебоидный тип организации характеризуется тем, что для передвижения и питания организмы изменяют форму поверхности тела, образуя цитоплазматические выступы – псевдоподии (ложноножки). Отсюда отсутствие плотных покровов тела, слабая обособленность эктоплазмы от эндоплазмы. В псевдоподиях особенно много микротрубочек. Система микротрубочек и микрофиламентов так же не позволяет перебалтываться внутреннему содержимому таких организмов. Средняя абсолютная скорость передвижения у простейших с амебоидным типом организации в 100–150 раз меньше, чем у жгутиконосцев. Зато их средние размеры больше в четыре раза (много полиэнергидных видов).
Возможно, что этот тип организации возник еще у прокариот, или как результат обитания в определенной среде (например, паразитические формы). Наиболее вероятным считают неоднократное возникновение амебоидных форм при переходе к обитанию на субстрате.
Плазмалемма простейших может содержать специальные рецепторные молекулы, позволяющие избирательно накапливать из среды различные вещества путем пиноцитоза. С внутренней стороны плазмалеммы обычно располагаются субмембранные микрофиламенты или микротрубочки. Совокупность их с плазмалеммой называется тубулеммой.
56
Энергетическое обеспечение метаболизма большинства видов простейших осуществляется с помощью митохондрий. Лишь у некоторых анаэробных свободноживущих и паразитических видов митохондрии отсутствуют. Первично лишенными митохондрий считают микроспоридий.
Специфическими органеллами простейших являются экструсомы. Известно 10 типов экструсом. Они выбрасываются наружу и служат для защиты или нападения.
В подцарство Protozoa объединены все наиболее примитивные эукариоты. Поэтому среди них наблюдаются все известные для эукариот типы питания, варианты строения клеток, возможные механизмы жизнедеятельности и жизненные циклы.
Разноклеточные. Строение на клеточном и надклеточном уровнях. Ткани и органы. Макротаксономия и краткая характеристика основных таксонов – растений, грибов и животных
Высшие эукариотические организмы (растения, грибы, животные) часто называют многоклеточными. Но многоклеточные организмы есть и среди простейших. Поэтому термин гетероклеточные (или разноклеточные) представляется более соответствующим действительному строению этих организмов. Ведь они, в отличие от многоклеточных простейших, состоят из разных по строению и специализированных по выполняемым функциям клеток. Изучает строение и функции клеток наука цитология.
Клетки (эуциты) всех трех основных групп организмов – растений (рис. 7), животных (рис. 8) и грибов (рис. 9) – являются эукариотными. Их ДНК (у человека молекула ДНК состоит из примерно 3·109 пар нуклеотидов), связанная с белком, организована в хромосомы, которые располагаются в особом, самом крупном органоиде клетки – ядре (лат. – nucleus; греч. – karyon). Ядро окружено ядерной оболочкой (кариолемма) из двух мембран. Она пронизана довольно крупными порами, через которые могут переходить из ядра в цитоплазму и обратно достаточно крупные частицы, например, субчастицы рибосом. Наружная мембрана ядра в некоторых местах объединяется с эндоплазматической сетью и может быть усеяна рибосомами. Ядро – это место хранения и воспроизведения наследственной информации. В тесном взаимодействии с цитоплазмой оно служит центром управления обменом веществ и почти всех процессов, происходящих в клетке. В норме ядро отсутствует лишь у эритроцитов и в зрелых члениках ситовидных трубок флоэмы. Чаще всего в клетке имеется одно ядро, редко – два или несколько (многоядерны поперечнополосатые
57
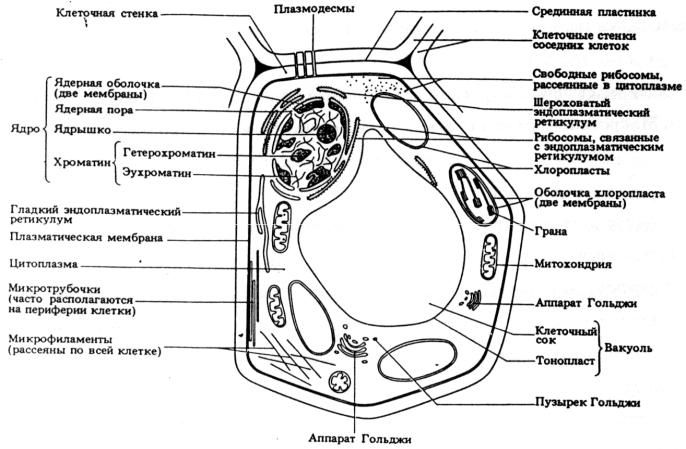
Рис. 7. Ультраструктура обобщенной растительной клетки, выявляемая при помощи электронного микроскопа (из Грин, Стаут, Тейлор, 1996)
58
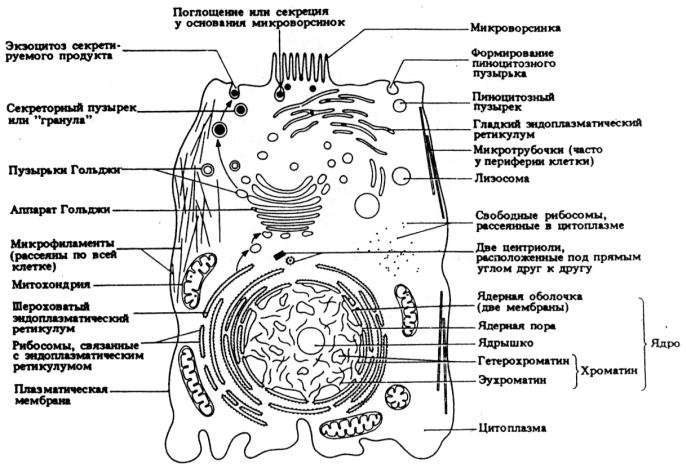
Рис. 8. Ультраструктура обобщенной животной клетки, выявляемая при помощи электронного микроскопа (из Грин, Стаут, Тейлор, 1996)
59
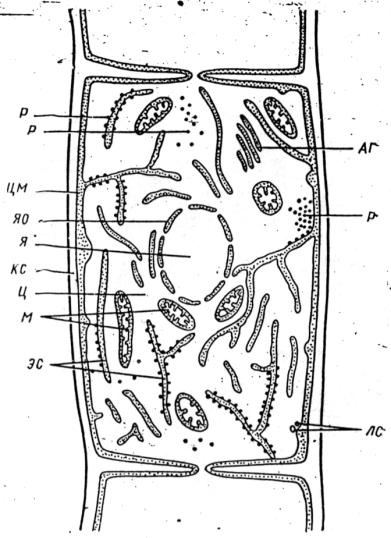
Рис. 9. Схема строения грибной клетки (из Великанов и др., 1981):
кс – клеточная стенка, я – ядро, яо – ядерная оболочка, р – рибосомы, м – митохондрии, цм – цитоплазматическая мембрана, лс – ломасомы, аг – аппарат Гольджи (диктиосомы), эс – эндоплазматическая сеть, ц – цитоплазма
60
мышечные волокна). Длительное существование в одной клетке двух не слившихся ядер (дикарион) обычно для некоторых грибов. Размеры ядер от 2–3 до 1000 мкм (в яйцах некоторых рыб и земноводных). Форма обычно шаровидная или эллипсоидная. Содержимое ядра представляет собой гелеобразный матрикс, называемый нуклеоплазмой (или ядерным соком). Внутри ядра обычно имеется одно, реже несколько ядрышек. Ядрышко или нуклеола – плотное тельце без мембраны, свободно лежащее в кариоплазме. Состоит в основном из белка и примерно 5 % РНК. Основная функция – синтез РНК (в основном рибосомной РНК) и формирование предшественников рибосом. При митозе ядрышко обычно распадается, а по его окончании формируется заново.
Живое содержимое клеток исторически принято называть протоплазмой. Клетка лишенная клеточной стенки и имеющая только клеточную мембрану, которая ограничивает снаружи цитоплазму с органеллами, называется протопластом. Есть другое определение протопласта – это активное содержимое клетки, отграниченное от клеточной стенки плазмалеммой, а от вакуоли тонопластом. Клетки животных, растений и грибов имеют много общего в своем строении. Начнем с одинаковых черт строения.
Цитоплазма – обязательная часть клетки, заключенная между плазмалеммой и ядром. Основу цитоплазмы составляет ее матрикс или гиалоплазма (от греч. hyalos – стекло и plasma – вылепленное, оформленное; плазмой называют жидкую или гелеобразную часть биологических структур). Это сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. В ее состав помимо воды (ее доля составляет около 90 %) входят растворимые белки, растворимые РНК, полисахариды, липиды (растворимую часть цитоплазмы называют цитозоль). Гиалоплазма объединяет все клеточные структуры в единую систему и обеспечивает взаимодействие между ними в процессах клеточного метаболизма. Именно в цитоплазме происходит анаэробный процесс расщепления углеводов с высвобождением энергии – гликолиз. Цитоплазма пронизана микротрубочками, филаментами и микрофиламентами. Совокупность таких тонких тяжей составляет цитоскелет (микротрабекулярная решетка). В животных клетках выделяют два слоя цитоплазмы. Наружный слой – эктоплазма (лишена гранул и большинства органоидов, обладает относительно высокой вязкостью, в ней есть сплетение микрофиламентов). Внутренний слой – эндоплазма (содержит гранулы и органоиды, обладает меньшей вязкостью). В живых клетках бывает заметно активное движение цитоплазмы – ток цитоплазмы или циклоз.
61
Помимо ядра и цитоплазмы все типичные клетки имеют клеточные мембраны. Они отделяют содержимое клетки от внешней среды (плазмалемма) и делят его на компартменты (отсеки), предназначенные для тех или иных метаболических путей. Благодаря избирательной проницаемости клеточные мембраны регулируют обмен между клеткой и средой. На мембранах располагаются рецепторные участки для распознавания внешних стимулов. Некоторые химические реакции протекают на мембранах (например, окислительное фосфорилирование при дыхании в митохондриях).
Структурную основу мембран составляют молекулы липидов. Они располагаются упорядоченно в два параллельных слоя. Гидрофильные части молекул липидов направлены наружу, а гидрофобные остатки жирных кислот — внутрь. Разными способами в мембрану встроены молекулы белков, большинство из которых является ферментами. Следовательно, мембраны – это липопротеиновые структуры. На внешней поверхности к некоторым липидным и белковым молекулам присоединены углеводные компоненты (гликозильные группы). На долю углеводов в мембранах приходится от 2 до 10 %. В большинстве случаев толщина мембран составляет 4–10 нм.
Внутри цитоплазмы мембраны образуют эндоплазматическую сеть или ретикулум (от лат. reticulum – сеть) – трехмерную систему цистерн (мембранных мешочков) и канальцев, соединенных друг с другом. Ее строение зависит от метаболической активности, стадии дифференцировки и типа клетки. Сеть, покрытая рибосомами или их группами (полисомами), получила название шероховатой. Если рибосомы отсутствуют, то это гладкий эндоплазматический ретикулум (гладкий ЭР). Функция шероховатого ЭР – транспорт белков, синтезируемых рибосомами на его поверхности. Гладкий ЭР секретирует липиды.
Комплекс (или аппарат) Гольджи впервые обнаружил в 1898 г. итальянский ученый Камилло Гольджи. Эта структура представляет собой стопку цистерн, и связанную с ними систему пузырьков (пузырьки Гольджи). В растительных клетках присутствует ряд отдельных стопок – диктиосом. В клетках животных чаще встречается одна большая стопка. Комплекс Гольджи секретирует и модифицирует клеточные продукты. Например, считают, что в диктиосомах образуются полисахариды. Аппарат Гольджи бокаловидных клеток эпителия слизистой оболочки кишечника и дыхательных путей секретирует гликопротеин муцин, в растворе образующий слизь. Транспорт продуктов осуществляют пузырьки Гольджи.
Рибосомы – очень мелкие органеллы (диаметром от 17 до 23 нм). Рибосомы эукариот (80S-рибосомы, лишь в митохондриях и пластидах 70S-рибосомы) состоят из двух не-
62
равных по величине субъединиц. В состав субъединиц входят рибосомная РНК (рРНК) и белок. Рибосомная РНК синтезируется в ядрышке. Рибосомы (в клетке их может быть несколько десятков тысяч) свободно располагаются в цитоплазме или прикреплены к эндоплазматической сети. Функция рибосом (вернее полисом) – синтез белка.
Митохондрии (от греч. mitos – нить и chondrion – зернышко, крупинка) – органоиды всех аэробных клеток, обеспечивающие организм энергией. Описаны Р.Альтманом в 1894 г. под названием биобластов. Митохондриями их назвал в 1897 г. К.Бенда. Форма (чаще эллиптические или округлые), количество (от нескольких десятков до 100 тыс.) и величина митохондрий постоянно меняются. Средний размер 0,2 – 1мкм. Митохондрия состоит из матрикса, окруженного внутренней мембраной (образует выросты в полость митохондрии
– кристы), межмембранного пространства и двух наружных мембран. В матриксе содержится собственная митохондриальная ДНК и рибосомы прокариотного типа. Митохондрии могут независимо от ядра синтезировать свои белки. Образуются митохондрии только в результате деления путем перешнуровки. Основная функция митохондрий – образование энергии (до 95 % в животной клетке и немного меньше в растительной и клетке грибов), путем дыхания. Богатые энергией молекулы АТФ синтезируются при реакции окислительного фосфорилирования. Энергия получается в результате окисления различных веществ, главным образом сахаров. Существует гипотеза, о том, что такие, в известной степени, автономные, органеллы как митохондрии и пластиды представляют собой видоизмененные прокариотические организмы, приобретенные гетеротрофными клетками-хозяевами в результате независимых случаев внутреннего симбиоза (эндосимбиоза).
Пероксисомы или микротельца – обычные органеллы большинства типов клеток. Имеют округлую форму. Диаметр 0,3–1,5 мкм. Они окружены одинарной мембраной. В тонкогранулярном содержимом пероксисом иногда видно кристаллическое ядро (кристаллический белок (фермент)). Их происхождение связано с деятельностью эндоплазматического ретикулума. Содержат фермент каталазу, катализирующий разложение токсичного пероксида водорода (побочный продукт некоторых окислительных процессов, протекающих в клетке) на воду и кислород. Очень много пероксисом в клетках печени. Пероксисомы животных клеток принимают участие в некоторых метаболических процессах, связанных с окислением. В растительных клетках выделяют три группы пероксисом. Глиоксисомы (имеют отношение к метаболизму глиоксилата) участвуют в превращении липидов в сахарозу в некоторых богатых маслами семенах (например, в эндосперме семян клещевины). Пероксисомы листьев играют важную роль в фотодыхании, так как связаны в нем с
63
хлоропластами и митохондриями (пероксид водорода один из продуктов фотодыхания). В других тканях растений найдены неспециализированные пероксисомы.
Микротрубочки – полые цилиндрические неразветвленные органеллы. Диаметр около 24 нм. Толщина стенок около 5 нм. Длина до нескольких микрометров. Построены из спирально упакованных глобулярных субъединиц белка тубулина. Растут с одного конца путем добавления тубулиновых субъединиц. От стенок через определенные промежутки отходят выступы, образующие связи с соседними микротрубочками. Из них состоит опорная система клетки – цитоскелет. Микротрубочки участвуют в перемещении клеточных органелл, например, пузырьков Гольджи. Нити веретена, появляющегося при делении ядра, также построены из микротрубочек.
Микрофиламенты – очень тонкие белковые (белок актин) нити диаметром 5–7 нм. Микрофиламенты образуют цитоскелет сходный с цитоскелетом из микротрубочек. Микрофиламенты образуют сплетения и пучки непосредственно под плазмалеммой. Они способны к сокращению. Участвуют в изменении формы клеток. Вероятно, микрофиламенты ответственны за циклоз в растительных клетках.
Вакуоли (франц. vacuole от лат. vacuus – пустой) – полости в цитоплазме, ограниченные мембраной и заполненные жидкостью. У простейших есть пищеварительные и сократительные вакуоли. Для гетероклеточных животных характерны пищеварительные и аутофагирующие вакуоли, входящие в группу вторичных лизосом (содержат гидролитические ферменты). У растений вакуоли – производные эндоплазматической сети, ограниченные мембраной – тонопластом и заполненные клеточным соком. В молодых клетках растений вакуоли представляют систему канальцев и пузырьков (провакуоли). По мере роста клеток они увеличиваются и сливаются в одну большую центральную вакуоль (занимает от 70 до 90 % объема клетки). Клеточный сок – слабокислый (pH 2–5) водный раствор различных органических и неорганических веществ.
Включения – относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (гликоген в клетках животных и большинства грибов, крахмал в клетках растений), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты).
Есть и существенные, специфические черты, свойственные клеткам каждой из групп организмов. Так, клетки растений и грибов, как и у большинства прокариот, имеют кнаружи от плазмалеммы клеточную стенку (она же клеточная оболочка). Клеточная стенка растений это структурное образование, придающее клетке прочность, сохраняющее ее
64
форму и защищающее протопласт. Она противостоит осмотическому давлению большой центральной вакуоли и препятствует разрыву клетки. Совокупность клеточных стенок играет роль внешнего скелета, поддерживающего форму растения и придающего механическую прочность. Клеточная стенка способна к росту растяжением. Она прозрачна и хорошо пропускает солнечный свет. Через нее легко проникают вода и низкомолекулярные вещества. Клеточные стенки соседних клеток скреплены между собой пектиновыми веществами, образующими срединную пластинку. Основу клеточной стенки растений составляют высокополимерные углеводы: молекулы целлюлозы (клетчатки), собранные в сложные пучки – фибриллы (или микрофибриллы), образующие каркас, погруженный в основу (матрикс), состоящий из гемицеллюлоз, пектинов и гликопротеидов. Молекулы целлюлозы (не растворяются в разбавленных кислотах и концентрированных щелочах) состоят из большого числа мономеров – остатков глюкозы. Клеточные стенки сильно гидратированы: 60–70 % их массы обычно составляет вода. Материалы, состоящие из более чем одного компонента и имеющие повышенную механическую прочность, называют композиционными материалами (композитами). Оболочки делящихся и растущих клеток называют первичными (они богаты водой и содержат мало целлюлозы – не более 30 %). Тонкие участки первичной оболочки называются поровыми полями. У некоторых клеток процесс отложения оболочки изнутри может продолжаться (вторичное утолщение оболочки). Такая оболочка называется вторичной. Она выполняет главным образом механическую функцию (содержит 40–50 % целлюлозы от массы сухого вещества). В стенках соседних клеток могут образовываться поры (обычно там, где есть поровые поля). Поры – это отверстия во вторичной оболочке, где клетки разделяет только первичная оболочка и срединная пластинка. Такие участки называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку пронизывают плазмодесменные канальцы (сквозного отверстия обычно не образуется). Содержимое соседних клеток связано друг с другом цитоплазматическими тяжами – плазмодесмами. Единая система протопластов называется симпластом (синцитием). Посредством плазмодесм передается раздражение и возможно передвижение некоторых веществ. Некоторые клетки претерпевают одревеснение – лигнификацию. При этом все слои целлюлозы пропитываются лигнином – полимерным веществом полифенольной природы. Стенка, пропитанная лигнином, становится прочной и твердой. Стенки клеток могут включать слои липидов: воска, кутина и суберина. Воск и кутин обычно покрывают наружные стенки клеток эпидермы. Так создается водо- и воздухонепроницаемый слой. Суберин пропитывает стенки. Так как он непроницаем для воды и газов, то клетки отми-
65
рают, и образуется пробковый слой. Клеточные стенки грибов тонкие (обычно толщина около 0,2 мкм), жесткие. Основной компонент хитин – азотсодержащий полисахарид, формирующий фибриллы. Иногда (отдел Oomycota) клеточная стенка содержит целлюлозу. У многих дрожжей скелетная часть стенки состоит из глюканов, отличных от целлюлозы. В клеточной стенке большинства видов грибов хорошо различимы слои. Наружный слой часто аморфный, а внутренний представляет гомогенный матрикс, с погруженными в него и определенным образом ориентированными микрофибриллами. Иногда клеточная стенка имеет еще более сложное строение. Так, у нейроспоры (Neurospora) она состоит из четырех хорошо дифференцированных слоев. Аморфный матрикс образован преимущественно глюканами с разными типами связей.
Только для клеток растений (и некоторых фотосинтезирующих простейших) характерны пластиды. У высших растений они образуются из пропластид – мелких обычно бесцветных телец, обнаруживаемых в меристематических зонах растения. Если из-за отсутствия света развитие пропластид в более дифференцированные структуры задерживается, то в них появляется одно или несколько проламеллярных телец (скопления трубчатых мембран). Такие бесцветные пластиды называются этиопласты. На свету они превращаются в хлоропласты. Различают три основных типа пластид: хлоропласты (зеленого цвета), хромопласты (желтого, оранжевого или красного цвета) и лейкопласты (бесцветные). В клетке обычно встречаются пластиды только одного типа. Известно, что одни типы пластид могут переходить в другие. Хлоропласты могут достигать в длину 4–10 мкм. В клетке может быть несколько десятков пластид. Форма окрашенных пластид обычно линзовидная или эллиптическая. У водорослей пластиды часто крупнее (количество обычно от 1 до 5) и разнообразнее по форме. Их называют хроматофорами. Хлоропласты имеют оболочку из двух мембран: наружной и внутренней. Внутренняя мембрана вдается в полость хлоропласта немногочисленными выростами. Оболочка отграничивает от гиалоплазмы клетки матрикс хлоропласта – строму. Строма и выросты внутренней мембраны формируют в полости хлоропласта сложную систему мембранных поверхностей, отграничивающих плоские мешки, называемые тилакоидами или ламеллами. Группы дисковидных тилакоидов связаны друг с другом таким образом, что их полости оказываются непрерывными. Эти тилакоиды образуют стопки – граны. Тилакоиды стромы объединяют граны между собой. В мембранах тилакоидов находится хлорофилл (несколько его модификаций) и вспомогательные пигменты – каротиноиды. Внутренняя структура хромопластов и лейкопластов проще – нет гран. В строме хлоропластов содержатся ферменты и рибосомы прокариотно-
66
го типа. Иногда имеются зерна первичного ассимиляционного крахмала. Есть собственная ДНК (могут синтезировать собственные молекулы белка). Основная функция хлоропластов – фотосинтез. Световые реакции фотосинтеза осуществляются преимущественно в гранах, а темновые в строме. В хлоропластах осуществляется так же синтез АТФ, АДФ, ассимиляционного крахмала и синтез и гидролиз липидов. Хромопласты считают конечным этапом в развитии пластид. Они определяют яркую окраску цветков, плодов и осенних листьев. В лейкопластах может осуществляться синтез и накопление запасных питательных веществ (крахмал (амилопласты), реже белки (протеинопласты) и жиры (липи-
допласты, элайопласты или олеопласты)).
В клетках животных и грибов имеются лизосомы (от греч. lysis – разложение, распад, растворение и soma – тело) – органеллы в виде пузырьков диаметром 0,2–0,5 мкм, окруженных одинарной мембраной. Образуются в комплексе Гольджи. Содержат набор гидролитических ферментов. С их помощью осуществляется внутриклеточное пищеварение, автофагия (переваривание частей самой клетки) и автолиз – саморазрушение клетки (например, при резорбции хвоста у головастика или после гибели клетки).
Центриоли (от лат. centrum – срединная точка, центр) – органоиды, встречающиеся почти во всех животных клетках и клетках низших растений. Впервые описаны В.Флеммингом в 1875 г. Это мелкие полые цилиндры. Располагаются парами в характерно окрашиваемой области цитоплазмы, известной под названием центросома или центросфера. Каждая центриоль построена из девяти триплетов микротрубочек. В начале деления ядра клетки, центриоли удваиваются, и уже две пары центриолей расходятся к полюсам веретена – структуры, по экватору которой выстраиваются перед своим расхождением хромосомы. Центриолям по структуре идентичны базальные тельца (кинетосомы или блефаропласты). Они обнаруживаются в основании жгутиков и ресничек.
Десмосомы – специализированные контактные участки (0,2×0,5 мкм) между животными клетками. Наиболее распространены в эпителиальных тканях. Плазмалеммы двух контактирующих клеток в десмосоме идут параллельно друг другу и разделены пространством шириной около 30 нм, в котором располагается тонкая пластинка плотного вещества. Иногда межмембранное пространство пронизано поперечными перегородками, соединяющими две мембраны (перегородчатые или сотовидные десмосомы у беспозвоночных). К внутреннему слою каждой мембраны в десмосоме прилегает электронноплотное вещество толщиной около 0,1 мкм, в которое, описывая дугу, входят из цитоплазмы филаменты из прекератина.
67
Ломасомы – мембранные структуры грибных клеток, имеющие вид многочисленных пузырьков. Располагаются между клеточной стенкой и плазмалеммой. По происхождению различают настоящие ломасомы и плазмалеммасомы.
Клетки позвоночных животных в состоянии жить и избегать высыхания, только находясь в жидкой среде. Такой средой им служит тканевая (интерстициальная или межклеточная) жидкость, содержащая помимо воды довольно значительное количество растворенных веществ.
Помимо собственно клеток (эуцитов) гетероклеточные организмы могут состоять из сложных цитоидов: плазмодиев, симпластов и синцитиев (сомателла простейших тоже сложный цитоид). Диаметр большинства клеток эукариот лежит в пределах 10–100 мкм, но может быть и значительно больше в случае некоторых специализированных клеток. Так одноклеточные волоски семян некоторых сортов хлопчатника достигают 5 см длины, а одноклеточные волокна растения рами даже 55 см (поперечник этих клеток всего 50–100 мкм).
Существование клетки от ее образования до деления на дочерние (или смерти) называют жизненным или клеточным циклом. У одноклеточных простейших он совпадает с жизнью особи. В непрерывно размножающихся тканевых клетках жизненный цикл совпадает с митотическим циклом.
Как отметил Л.Н.Серавин многоклеточные (и полицитоидные) организмы в процессе эволюции возникали в разных ветвях филогенетического дерева не менее 18 раз. Р.Н.Буруковский полагает, что это вполне объяснимый закономерный процесс. По современным представлениям материя характеризуется тенденцией к самоорганизации, к возникновению порядка из хаоса (взгляды И.Пригожина). В природе наблюдается тенденция к возникновению все более сложных систем. Они появляются на разных уровнях организации материи закономерно всякий раз, когда для этого складываются необходимые условия. Поэтому многоклеточность не могла не возникнуть у простейших, растений и животных. В трех линиях растительных (Rhodophyta – красные водоросли, Pheophyta – бурые водоросли и Plantae – высшие растения) и трех линиях гетеротрофных организмов (миксомомицеты класса Dictyostelida, Metazoa и губки – Spongia) эволюция пошла по пути возникновения гетероклеточности, связанной с дифференцировкой соматических клеток.
Дифференцировка – это процесс, в результате которого клетка (вернее группа клеток) приобретает морфологические, химические и функциональные особенности в результате разделения физиологического труда, т.е. становится специализированной. Например,
68
в организме человека насчитывают около 100 различных типов клеток: клетки пищеварительного тракта синтезируют пепсин и трипсин, фибробласты – коллаген, миобласты – миозин, нейроны принимают сигналы, перерабатывают их в нервные импульсы и проводят к нервным окончаниям и т.д. Разнообразие эукариотических клеток показано на рис. 10. Дифференцировка лежит в основе морфогенеза и происходит в основном в процессе зародышевого развития, а также в некоторых органах взрослого организма. Так, в кроветворных органах тотипотентные (тотипотентность – способность клеток реализовывать генетическую информацию ядра, обеспечивающую их дифференцировку) стволовые кроветворные клетки дифференцируются в различные клетки крови. Поэтому в более широком смысле под дифференцировкой понимают постепенное (на протяжении нескольких клеточных циклов) возникновение все больших различий и направлений специализации между клетками, произошедшими от клеток одного исходного зачатка. Считается, что дифференцировка клеток в процессе онтогенеза является результатом последовательных реципрокных (от лат. reciprocus – взаимный) влияний цитоплазмы и меняющихся продуктов активности ядерных генов. В большинстве случаев соматические клетки организмов несут полный диплоидный набор хромосом и их ядра полностью сохраняют генетические потенции. Об этом свидетельствуют опыты Дж.Гердона по пересадке в энуклеированную ультрафиолетовым облучением яйцеклетку шпорцевой лягушки Xenopus laevis ядра из эпителиальной клетки кожи лягушки или кишечника головастика. Из таких яйцеклеток развивались нормальные особи.
Системы специализированных клеток, сходных по происхождению, строению и функциям, включающие также некоторые продукты их жизнедеятельности (межклеточные вещества и структуры), образуют ткани (лат. – textus, греч. – histos). Изучает ткани наука гистология.
Простые ткани состоят из одного типа клеток. У животных простой тканью является плоский эпителий, у растений – паренхима и колленхима. Сложные ткани содержат клетки разных типов. К ним у растений относят ксилему и флоэму, а у животных рыхлую (ареолярную) соединительную ткань.
Развитие высших растений начинается со стадии зародыша. Большинство его клеток обладает меристематической активностью (способностью к делению). Часть из них сохраняет эту способность в течение длительного времени. Это инициальные клетки или инициали. Клетки, постоянно возникающие при делении инициалей, называют производными. Они делятся один или два раза и начинают дифференцироваться в специализированные
69
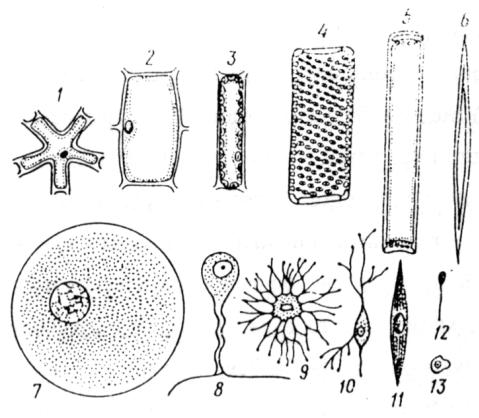
Рис. 10. Разнообразие форм эукариотических клеток (из Мамонтов, Захаров, 1999): Растительные клетки: 1, 2 – паренхимные, 3 – из палисадной ткани, 4 – клетки сосуда, 5 – клетка ситовидной трубки, 6 – клетка механической ткани. Клетки животных: 7 –
яйцеклетка, 8 и 10 – нервные клетки, 9 – клетка костной ткани (остеоцит), 11 – клетка гладкой мышцы, 12 – сперматозоид, 13 – лейкоцит
70
клетки. Образуются постоянные ткани, клетки которых лишены способности к делению. Лишь в некоторых случаях, когда специализация не зашла слишком далеко, клетки постоянных тканей могут вновь приобретать меристематическую активность.
Одна из особенностей растений – способность к длительному росту – определяется наличием у них образовательных тканей (меристем), которые формируют все прочие ткани за счет деления и образования новых клеток. Существует два основных типа мери-
стем – апикальные (верхушечные) и латеральные (боковые). Апикальные меристемы рас-
полагаются на верхушках корней и побегов и обеспечивают их нарастание в длину (первичный рост, соответственно это первичные меристемы). К первичным меристемам относят и производные апикальных меристем, сохраняющие в определенной степени способ-
ность к делению. Это протодерма, прокамбий и основная меристема. Еще в ходе эмбрио-
генеза они дают три первичные системы тканей: протодерма – покровную ткань, прокамбий – проводящую и основная меристема – систему основных тканей (основная ткань – паренхима). Первичны интеркалярные (вставочные) и раневые меристемы. Интеркалярные меристемы сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлий или в основаниях черешков листьев злаков). Раневые меристемы образуются в местах повреждения тканей и органов и дают начало каллусу – особой ткани, состоящей из однородных паренхимных клеток. Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов. Соответственно они обычно образуют цилиндры, которые на поперечных срезах имеют вид колец. Главные латеральные меристемы – камбий и феллоген. Они обеспечивают нарастание стволов в толщину, образуя вторичные ткани. Так камбий дает начало вторичным проводящим тканям – вторичной ксилеме и флоэме, а феллоген – пробке.
Внутренние ткани растений от повреждений, проникновения патогенных организмов и высыхания защищают покровные ткани. Они также регулируют газообмен и транспирацию – испарение воды растением (от лат. trans – через и spiro – дышу, выдыхаю). Некоторые покровные ткани способны к всасыванию и выделению, активно регулируя скорость и избирательность проникновения веществ. Большинство покровных тканей состоит из плотно сомкнутых живых, реже мертвых клеток. К первичным покровным тканям относятся эпидерма (чаще однослойная, на листьях и зеленых побегах). Транспирация и газообмен происходят через устьица. Они состоят из двух замыкающих клеток, между которыми имеется устьичная щель. Щель может расширяться и сужаться. В движении замыкающих клеток принимают участие примыкающие к ним побочные клетки. Вместе они
71
образуют устьичный аппарат. Изучает строение устьичных аппаратов растений стоматография. Полученная информация используется в систематике растений. На эпидерме могут находиться выросты: трихомы (кроющие и железистые), эмергенцы (жгучие волоски крапивы, шипы роз и т.д.). Первичная однослойная покровная ткань корня – эпиблема (ризодерма). Покрывает молодые корневые окончания. Через нее происходит поглощение воды и минеральных солей из почвы. Устьиц нет, зато есть корневые волоски, которые чаще всего формируются из клеток – трихобластов.
Перидерма – сложная, многослойная, вторичная покровная ткань стеблей и корней многолетних (реже однолетних) растений. Сменяет отмирающие первичные покровные ткани. В ее основе лежит феллоген. В процессе формирования перидермы наружу откладываются клетки пробки, а внутрь живые паренхимные клетки феллодермы. Пробка, феллоген и феллодерма образуют перидерму (т.е. перидерму можно рассматривать как комплекс тканей). Для газообмена в перидерме закладываются чечевички – отверстия, прикрытые рыхлой тканью из закругленных, паренхимных слабоопробковевших клеток с многочисленными межклетниками. В многолетних осевых органах растений обычно развивается несколько перидерм. Каждая последующая – глубже предыдущей. Со временем клетки наружных перидерм (и тканей между ними) отмирают, образуя мощный трещиноватый и неровный защитный комплекс – корку (ритидом).
Большую часть тела растений составляют относительно мало специализированные основные ткани. Они располагаются между другими постоянными тканями во всех вегетативных и репродуктивных органах. Состоят обычно из живых паренхимных клеток, разнообразных по форме. Исходя из главной выполняемой функции, различают несколько групп основных тканей. Ассимиляционная ткань (хлоренхима) – состоит из клеток, содержащих хлоропласты. Эта ткань чаще всего залегает под эпидермой, особенно в листьях. Нередко, хлоренхима расположена очень рыхло, образуя крупные газоносные межклетники (т.е. добавляется воздухоносная функция). Избыточные в данный период развития растения продукты метаболизма откладываются в запасающих тканях, расположенных в различных органах (эндосперме или зародыше семян, клубнях, луковицах, сердцевине стеблей и др.). Запасающая ткань состоит из живых обычно тонкостенных клеток и может превращаться в хлоренхиму. Водоносная ткань служит для запасания воды. Крупноклеточная тонкостенная водоносная паренхима есть в листьях и стеблях суккулентов (кактусы, агавы, алоэ). Крупные водоносные клетки встречаются в листьях злаков. В вакуолях клеток водоносной паренхимы есть слизистые вещества, удерживающие влагу. Паренхиму с развитыми меж-
72
клетниками называют воздухоносной тканью или аэренхимой. Ее назначение – снабжение тканей кислородом, углекислым газом и обеспечение плавучести. Особенно она развита в органах болотных и водных растений.
Прочность органам растений придают механические ткани. Главная особенность клеток механических тканей – значительно утолщенные оболочки, которые продолжают выполнять опорные функции даже после отмирания их живого содержимого. Различают два основных типа механических тканей: колленхиму и склеренхиму.
Колленхима – простая первичная ткань, состоящая из вытянутых вдоль оси органа клеток с неравномерно утолщенными неодревесневшими первичными оболочками. По характеру утолщений стенок и соединению клеток между собою выделяют уголковую, пластинчатую и рыхлую колленхиму. Функцию арматурной ткани колленхима выполняет только в состоянии тургора (тургор – напряженное состояние клеточной оболочки, создаваемое гидростатическим давлением внутриклеточной жидкости). Колленхима обычно располагается под эпидермой, в черешках или по обеим сторонам крупных жилок. В корнях встречается редко. Клетки ее живые и способны к росту в длину, не препятствуя росту органов.
Склеренхимой называется механическая ткань, состоящая из клеток с одревесневшими и равномерно утолщенными оболочками, обладающими прочностью, близкой к прочности стали. На определенном этапе дифференциации протопласт отмирает, и склеренхимные клетки выполняют свои функции, будучи мертвыми. По происхождению склеренхима бывает первичной (из клеток основной меристемы, прокамбия или перицикла) и вторичной (из клеток камбия). Различают два основных типа склеренхимы – волокна и склереиды. Волокна – сильновытянутые прозенхимные клетки с заостренными концами. Волокна флоэмы (луба) называют лубяными. Волокна ксилемы (древесины) называют древесинными или волокнами либриформа. Склереиды – структурные элементы, обычно возникающие из клеток основной паренхимы в результате утолщения и лигнификации их оболочек. Они очень разнообразны по форме. Могут встречаться по одиночке или в виде скоплений в стеблях (хинное дерево), плодах (груша), листьях (камелия) и семенах (некоторые бобовые). Считается, что функция склереид заключается в противостоянии сдавливанию.
Проводящие ткани. Служат для передвижения по растению растворенных в воде питательных веществ. Возникли у растений как следствие приспособления к жизни в двух средах: почвенной и воздушной. От корня к листьям движется восходящий или транспира-
73
ционный ток водных растворов солей. Нисходящий или ассимиляционный ток органиче-
ских веществ идет от листьев к корням. Восходящий ток осуществляется по трахеальным элементам ксилемы, а нисходящий по ситовидным элементам флоэмы. Все проводящие ткани являются сложными, т.е. состоят из морфологически и функционально разнородных элементов. Ксилема и флоэма формируются из одной и той же меристемы и располагаются рядом. Во многих органах растений ксилема и флоэма объединены в виде, так называемых, проводящих пучков. Существуют первичные и вторичные проводящие ткани. Первичные закладываются в листьях, молодых корнях и побегах. Они дифференцируются из клеток прокамбия. Вторичные проводящие ткани, обычно более мощные, возникают из камбия.
Ксилема или древесина. Первичная и вторичная ксилемы состоят из клеток одних и тех же типов, но первичная ксилема не имеет сердцевинных лучей. Дальний транспорт веществ осуществляется по трахеидам и сосудам, а ближний по паренхимным элементам. Трахеиды – это мертвые проземхимные клетки длиной в среднем 1–4 мм. Их стенки одревесневают, утолщаются и несут простые или окаймленные поры, через которые происходит фильтрация растворов. Трахеиды есть у спорофитов всех высших растений, а у большинства хвощевидных, плауновидных, папоротниковидных и голосеменных они являются единственными проводящими элементами ксилемы. Сосуды – это полые трубки, состоящие из отдельных члеников, располагающихся друг над другом. Между члениками имеются разного типа сквозные отверстия – перфорации, благодаря чему ток жидкости осуществляется свободно. Наибольшего развития сосуды достигают у покрытосеменных растений. Сосуды и трахеиды в большинстве случаев имеют вторичные кольчатые и лестничные утолщения (кольчатые и лестничные сосуды и трахеиды). Эти трахеальные элементы распределяются в древесине различным образом. На поперечном срезе они могут образовывать кольца (кольцесосудистая древесина) или могут рассеиваться более или менее равномерно по всей ксилеме (рассеяннососудистая древесина). Ксилема включает так же лучевые элементы – клетки, образующие сердцевинные лучи. В лучах хвойных встречаются лучевые трахеиды. В ксилеме покрытосеменных содержатся также неодревесневшие живые паренхимные клетки – древесинная паренхима (место хранения запасных веществ). По сердцевинным лучам и древесинной паренхиме осуществляется ближний транспорт веществ.
Флоэма – сложная проводящая ткань, осуществляющая транспорт продуктов фотосинтеза от листьев к местам их использования или отложения. Первичная флоэма образуется из прокамбия, вторичная (луб) из камбия. В стеблях флоэма расположена обычно сна-
74
ружи от ксилемы, а в листьях она обращена к нижней стороне пластинки. У первичной флоэмы отсутствуют сердцевинные лучи. В состав флоэмы входят ситовидные и механические элементы, паренхимные клетки и элементы сердцевинных лучей. К ситовидным элементам относятся: ситовидные клетки (основной проводящий элемент флоэмы у всех групп высших растений, исключая покрытосеменные) и ситовидные трубки покрытосеменных (состоят из клеток – члеников длиной 150 – 300 мкм, поперечником 20 – 30 мкм с ситовидными полями на концах и клеток спутниц). Паренхимные элементы (лубяная паренхима) состоят из тонкостенных клеток. В них откладываются запасные питательные вещества и, отчасти, по ним осуществляется ближний транспорт веществ. Сердцевинные лучи тоже состоят из тонкостенных паренхимных клеток и осуществляют ближний транспорт ассимилянтов.
Проводящие пучки – обособленные тяжи проводящей системы. Первоначально возникают из камбия. Бывают полными – состоящими из ксилемы и флоэмы и неполными – только из ксилемы или флоэмы. У большинства двудольных и голосеменных пучки открытые. В них часть прокамбия сохраняется и превращается в камбий, а пучок способен к вторичному утолщению. В закрытом пучке однодольных прокамбий весь превращается в проводящие ткани. В зависимости от взаимного расположения флоэмы и ксилемы различают несколько типов пучков. Коллатеральные пучки (открытые и закрытые) – флоэма лежит по одну сторону от ксилемы. Биколлатеральные пучки – одна часть флоэмы располагается снаружи, а другая с внутренней стороны ксилемы. Концентрические пучки – флоэма окружает ксилему или наоборот.
Выделительные или секреторные ткани. Это структурные образования, способные активно выделять из растения или изолировать в его тканях продукты метаболизма и ка- пельно-жидкую воду. Различают ткани внутренней (выделяемые вещества остаются внутри растения) и наружной секреции. Клетки выделительных тканей по форме обычно паренхимные и тонкостенные. Считают, что синтез жидких секретов связан с деятельностью внутриклеточных мембран и комплекса Гольджи. От избыточной воды и солей растения освобождаются через гидатоды – системы клеток, выделяющие водно-солевые растворы из подходящего к ним небольшого проводящего пучка. Клетки-идиобласты содержат кристаллы оксалата кальция, смесь терпеноидов, таннины и слизи. Вместилища выделений (схизогенные и лизигенные) – полости различной формы, располагающиеся в толще других тканей. Смоляные ходы и эфирномасляные каналы образуются схизогенно, но более или менее вытянуты и могут ветвиться. Млечники – особый тип выделительной ткани. Они
75
пронизывают все растение. В вакуолях млечников находится млечный сок – латекс. К наружным выделительным тканям относятся железистые головчатые волоски (например, у герани), железки и нектарники (выделяют нектар).
У животных выделяют четыре основных типа тканей, соответствующие основным соматическим функциям организма: эпителий, соединительную, мышечную и нервную ткани.
Эпителиальная ткань представляет собой однослойные или многослойные пласты, покрывающие внутренние и внешние поверхности любого животного. Ее главная функция
– защита нижележащих тканей от повреждений. Она развивается из эктодермы (эпителий кожи и выстилка переднего и заднего отделов пищеварительного тракта) и из энтодермы (эпителий остальных участков пищеварительного тракта, печени и поджелудочной железы). Эпителиальные клетки удерживаются вместе веществом, содержащим гиалуроновую кислоту (кислый мукополисахарид). Нижний слой клеток лежит на базальной мембране, состоящей из переплетения коллагеновых волокон. Кислород и питательные вещества поступают к клеткам эпителия путем диффузии из лимфатических сосудов, разветвляющихся в примыкающих межклеточных пространствах. Свободная поверхность эпителия часто бывает высоко дифференцированной и выполняет, помимо защитной, множество различных функций: секреции и экскреции, всасывания и др. В зависимости от количества клеточных слоев и формы клеток эпителий делят на несколько типов: плоский, кубический, цилиндрический, мерцательный. Для многослойного эпителия характерно постоянное изнашивание и слущивание клеток, образующих поверхностный слой. Утраченные клетки замещаются снизу, где клетки эпителия образуют герминативный слой (в нем идет клеточное деление). Иногда клетки секретируют кутикулу, покрывающую эпителий более или менее плотным слоем. Железы почти всегда имеют эпителиальное происхождение.
Соединительная ткань – это главная опорная ткань организма. Она представляет собой сложную структуру, в состав которой входят разнообразные клетки, развивающиеся из мезенхимы, происходящей из мезодермы зародыша; волокна нескольких типов; жидкий или полужидкий аморфный матрикс из гиалуроновой кислоты, хондроитина, хондроитинсульфата и кератинсульфата. Существует несколько типов соединительной ткани.
Собственно соединительную ткань подразделяют на оформленную или ориентированную (сухожилия, фасции, склера глаза и др.) и неоформленную или диффузную (волокна соединены в пучки, расположенные неупорядоченно), в которой выделяют плотную (компактную) ткань (например, соединительнотканная основа кожи) и рыхлую ткань (напри-
76
мер, подкожная клетчатка). К соединительной ткани относится и жировая ткань. Она состоит из клеток, содержащих в цитоплазме жировые включения. Играет роль энергетического депо организма, предохраняет его от потери тепла.
К скелетным тканям относятся хрящ и кость. Хрящ – это твердая, но гибкая ткань, состоящая из клеток, погруженных в упругое вещество – хондрин (он отлагается хондробластами). Хондрин содержит многочисленные тонкие волокна, состоящие в основном из коллагена. В конечном счете, хондробласты оказываются заключенными в полости – лакуны. В этом состоянии их называют хондроцитами. Снаружи хрящ покрыт перихондрием или надхрящницей – плотной оболочкой, состоящей из клеток и волокон. Здесь формируются новые хондробласты. Основное вещество хряща упруго и способно демпфировать ударные нагрузки, возникающие между суставными поверхностями костей.
Кость – это основной материал скелета позвоночных животных. Представляет собой обызвествленную соединительную ткань, состоящую из клеток, погруженных в твердое основное вещество. Его состав: 30 % органические соединения (коллагеновые волокна) и 70 % неорганические соединения (главный компонент гидроксиапатит – Ca10(PO4)6(OH)2 плюс натрий, магний, калий и др.). Костные клетки (остеобласты) находятся в лакунах, распределенных по всему основному веществу и соединенных тонкими канальцами с цитоплазмой. Остеобласты откладывают неорганическое вещество кости. Снаружи кость покрыта надкостницей.
Гематопоэтические ткани – миелоидная и лимфоидная. В миелоидной ткани (костный мозг) образуются эритроциты и гранулоциты. В лимфоидной ткани образуются лимфоциты и моноциты. Эти ткани состоят из свободных клеток, лежащих в строме, образованной рыхлыми ретикулиновыми волокнами (ретикулярная соединительная ткань).
Кровь – циркулирующая в кровеносной системе «жидкая ткань». Одна из форм соединительной ткани. Обеспечивает жизнедеятельность других тканей. Выполняет множество функций: дыхательную, трофическую, экскреторную, регуляторную, защитную. У позвоночных представляет собой однородную густую красную жидкость, состоящую из плазмы и форменных элементов (эритроцитов, лейкоцитов, тромбоцитов).
Мышечная ткань составляет основную массу мышц и осуществляет их сократительную функцию. Она образуется из мезодермы зародыша. Состоит из сократительных клеток или волокон, соединенных между собой соединительной тканью. По характеру ин-
нервации различают произвольные (поперечно-полосатые), непроизвольные (гладкие) мышцы и отдельно выделяют сердечную мышцу.
77
Волокна гладких мышц устроены проще и по размеру они мельче. Гладкая мускулатура располагается главным образом в оболочке пищеварительного тракта. Гладкие мышечные волокна есть и в других местах: кровеносных и лимфатических сосудах, соединительной ткани кожи и других частей тела. Типичное гладкое мышечное волокно – это тонкая веретеновидная клетка длиной в среднем несколько десятых долей миллиметра с единственным, расположенным в центре ядром. В основном гладкомышечные клетки стимулируются автономными нервными волокнами.
Поперечно-полосатые мышцы образуют произвольную мускулатуру, формирующуюся в основном из миотомов зародыша. Обычно эти волокна скомпанованы в виде отдельных мышц, которые прикрепляются к скелетным элементам и двигают их. Поперечнополосатые волокна – это крупные, вытянутые в длину, цилиндрические клетки. Их длина от 1 мм до нескольких сантиметров (иногда более 30 см). Клетки содержат множество ядер (симпласт). Волокно покрыто тонкой оболочкой – сарколеммой, включающей в себя клеточную мембрану и базальную пластинку. Как и в гладких волокнах, внутри клетки содержат много плотно упакованных продольных фибрилл. Но есть отличие: каждая фибрилла состоит из чередующихся светлых и темных участков, располагающихся во всех фибриллах на одном уровне. В результате на волокне образуются поперечные полосы, давшие им название. Мышцы слагаются из параллельно расположенных волокон. Между ними находится связующая соединительная ткань. Она же образует оболочку мышцы – эпимизий. Каждое нервное волокно обычно на конце разделяется и образует с нескольки-
ми мышечными волокнами нервно-мышечные соединения (концевые двигательные пла-
стинки). В состоянии покоя сарколемма электрически поляризована. Нервный импульс приводит к выбросу из нервных окончаний ничтожного количества ацетилхолина, что вызывает мгновенную деполяризацию сарколеммы, а за счет этого – мышечное сокращение.
Сердечная мышца относится к особому типу. Она имеет клеточное строение и поперечную исчерченность миофибрилл. Вся мышца сердца представляет собой единую сеть разделяющихся и соединяющихся тяжей. Вставочные диски более или менее разделяют эту сеть на короткие отрезки. В эмбриогенезе эта мышца образуется из мезенхимы мезодермального происхождения. Ее гистологические особенности, вероятно, объясняются непрерывной активностью.
Нервная ткань – комплексы нервных и глиальных клеток, специфичных для животных организмов. Развивается из эктодермы зародыша. Состоит из связанных между собой нервных клеток – нейронов. Нейроны специализированы к проведению нервных импуль-
78
сов и восприимчивы к раздражению. Нейроны, передающие импульсы в центральную нервную систему, называют афферентными (сенсорными) нейронами. Нейроны, передающие импульсы от центральной нервной системы, называют эфферентными (моторными) нейронами. Нейрон состоит из тела клетки (диаметром 3 – 100 мкм) и цитоплазматических отростков. Отростки, проводящие импульсы к телу клетки, называют дендритами. Они короткие, относительно широкие и распадаются на тонкие терминальные ветви. Отростки, проводящие импульсы от тела клетки к другим клеткам или периферическим органам называют аксонами или нервными волокнами. Они тоньше дендритов, их длина может достигать нескольких метров. Дистальный участок аксона распадается на многочисленные веточки с вздутиями на концах. Он способен к нейросекреции. Дистальный участок аксона соединяется с соседними нейронами на участках, называемых синапсами, которые бывают возбудительные и тормозные. Нервные волокна могут быть миелинизированными (покрытыми миелиновой оболочкой, образуемой шванновскими клетками) и немиелинизированными. На основании числа и расположения отростков нейроны делят на уни-, псевдоуни-, би- и мультиполярные. Пучки нервных волокон, окруженные соединительнотканной оболочкой (эпиневрием), образуют нервы.
Ткани гетероклеточных существ образуют органы и системы органов. Органом принято называть часть организма, имеющую определенное строение и выполняющую определенные функции. По выполняемым функциям можно выделить различные органы. Например: репродуктивные (служат для воспроизводства), вегетативные (у растений; для поддержания жизни конкретной особи), дыхания (для газообмена), выделения и т.д.
Разноклеточные организмы представлены тремя таксонами: царство Растения (Vegetabilia или Plantae), царство Грибы (Mycetalia, Fungi, Mycophyta или Mycota) и царство Животные (Metazoa).
Царство Растения. В основном прикрепленные фотоавтотрофные организмы с «неограниченным» ростом. Первичные продуценты органических веществ. Тело с высоким отношением поверхности к объему, что способствует более эффективному улавливанию света и обмену веществ. Часто наблюдается ветвление. Клетки с плотной клеточной стенкой, состоящей преимущественно из целлюлозы. В клетках имеются особые органеллы – пластиды, в том числе хлоропласты, содержащие пигмент хлорофилл. Центриолей нет. Большая, постоянно существующая вакуоль, содержит клеточный сок. Нервная система отсутствует, раздражимость регулируется гормонами. Включает три подцарства, из которых собственно гетероклеточными организмами являются высшие растения. Багрянки
79
(Rhodobionta) – для представителей подцарства характерно наличие хлорофилла a и d и дополнительных фотосинтезирующих пигментов (фикоциан, фикоэритрин), сходных с пигментами цианобактерий. Запасное питательное вещество багрянковый крахмал, близкий к амилопектину и гликогену. Ундулиподии отсутствуют. Настоящие водоросли (Phycobionta) и Высшие растения (Embryobionta или Embryophyta) на разных стадиях жизнен-
ного цикла могут иметь ундулиподии. Для них характерен хлорофилл a и c или a и b. Запасное вещество чаще всего – крахмал. Спорангии и гаметангии высших растений многоклеточные либо сильно редуцированы. Зигота у них дает начало многоклеточному зародышу, который развивается в спорофит. Вегетативное тело высших растений дифференцировано на органы и ткани и обычно расчленено на корень, стебель и лист (поэтому эти растения часто называют листостебельными или побеговыми – Cormophyta или Cormobionta). Высшие растения хорошо приспособлены к жизни в воздушной наземной среде. Заселение ими (высшими споровыми растениями) суши произошло, вероятно, 415 – 430 млн. лет назад. Известно не менее 300000 ныне существующих видов.
Царство Грибы. Одна из самых больших (более 100000 описанных видов) и процветающих групп организмов. Ранее относили к низшим растениям. Грибы достаточно загадочная группа и сравнительно малоизученная. Древнейшие ископаемые, похожие на грибы, имеют возраст около 800 млн. лет. Бесспорные находки грибов датируются возрастом 450 – 500 млн. лет. В систематике грибов еще очень много спорных вопросов. Обычно выделяют четыре таксона, имеющие общее происхождение: зигомицеты, аскомицеты, базидиомицеты и дейтеромицеты. Их считают отделами (в таком случае название заканчивается на -mycota) или классами (-mycetes). Своеобразие грибов определяется сочетанием признаков, как растений, так и животных. Это гетеротрофные организмы, не способные передвигаться. Тело настоящих грибов состоит из массы тонких ветвящихся трубчатых нитей – гиф, с апикальным ростом. Вся масса гиф образует мицелий (грибницу). Гифа окружена тонкой жесткой стенкой, основным компонентом которой является хитин (иногда целлюлоза). Протоплазма гиф либо совсем не разделена (нечленистые, несептированные, асептированные или ценоцитные гифы), либо разделяется поперечными перегородками – септами (членистые или септированные гифы). Образование септ не связано с делением ядер. В центре септ, как правило, есть небольшое отверстие, через которое протоплазма может перетекать из одного компартмента в другой. Часть мицелия, пронизывающая субстрат, получила название субстратного, а часть, располагающаяся на поверхности – воздушного. На воздушном мицелии обычно образуются органы размножения. У дрожжей вегетативное
80
тело представлено одиночными почкующимися или делящимися клетками. Так как грибы
– гетеротрофы, то для питания им нужны органические источники углерода, а также источник азота (обычно аминокислоты). Сапрофитные грибы выделяют ферменты, которые воздействуют на субстрат и способствуют его частичному перевариванию вне грибной клетки. Образовавшийся полупереваренный материал грибы всасывают всей поверхностью путем диффузии. Если гриб способен секретировать ферменты трех основных классов (карбогидразы, липазы, протеазы), то он способен использовать разнообразные субстраты и становится вездесущим, как, например Penicillium (к тому же грибы-сапрофиты обычно образуют большое количество легких устойчивых спор). Грибы-сапрофиты и бактерии образуют вместе группу организмов-редуцентов. Грибы-паразиты могут быть факультативными (часто вызывают гибель своих хозяев и потом живут сапрофитно на мертвых остатках) и облигатными (паразитируют на узком круге хозяев, от которых им нужен специфический набор питательных веществ). Облигатные паразиты для проникновения в клетки растений-хозяев, и высасывания из них питательных веществ, образуют специальные выросты – гаустории. Грибы участвуют в создании двух типов симбиозов. Симбиоз сумчатого или базидиального гриба и водоросли (обычно зеленые или реже сине-зеленые водоросли) называется лишайником. О характере взаимоотношений мико- и фикобионта единого мнения нет. Часто считают, что это гармоничный взаимовыгодный симбиоз. Однако многие специалисты склоняются к мнению, что имеет место контролируемый паразитизм гриба на автотрофе. Симбиоз гриба с корнями растений – микориза. При этом гриб получает от растения углеводы и витамины и расщепляет белки до аминокислот, часть которых поглощается растением.
Царство животные. Гетеротрофные подвижные организмы с активным метаболизмом и ограниченным ростом тела. Помимо гормонов имеют нервную систему, позволяющую реагировать быстро. У большинства есть специальные органы выделения, в частности для выведения азотистых веществ. Клетки не имеют жесткой клеточной стенки. Нет пластид. Вакуоли небольшие и недолгоживущие. Есть центриоли. Запасное вещество – гликоген. Царство Metazoa включает два подцарства: Prometazoa и Eumetazoa. Примитивные беспозвоночные животные из подцарства Prometazoa (губки, ортонектиды, дициемиды и единственный представитель типа Placozoa – трихоплакс) имеют паренхимелльный план организации. У них нет рта, кишечника, нервной системы и мускулатуры. Основной способ питания фагоцитоз и пиноцитоз. У настоящих многоклеточных (гетероклеточных) животных, объединенных в подцарство Eumetazoa, есть рот, как правило, кишечник, нервная
81
система и мускулатура. Они макрофаги. Происходят, по-видимому, от паренхимулообразных предков. В настоящее время выделяют около 20 типов многоклеточных животных. Многоклеточные животные появились, вероятно, 700 – 900 млн. лет назад. Первые ископаемые следы жизнедеятельности червеобразных многоклеточных животных известны из позднерифейских отложений. В вендское время (650 – 570 млн. лет назад) существовали уже разнообразные животные, принадлежавшие к различным типам.
82