

10.5. Анаэробный гликолиз
Анаэробный гликолиз - сложный ферментативный процесс последовательных превращений глюкозы, протекающий в тканях человека и животных без потребления кислорода (рис.28).
Обратимое превращение пировиноградной кислоты в молочную катализируется лактатдегидрогеназой:
![]()
-
Пировиноградная кислота
Молочная кислота
Суммарный результат гликолиза выражается следующим уравнением: С6Н12О6 + 2Н3РО4 + 2АДФ = 2С3Н6О3 + 2АТФ + 2Н2О
-
Глюкоза
Молочная кислота
Таким образом, чистый выход АТФ при анаэробном гликолизе - 2 моль АТФ на 1 моль глюкозы. Именно благодаря анаэробному гликолизу организм человека и животных может определенный период времени осуществлять ряд физиологических функций в условиях недостаточности кислорода.
Данный процесс у бактерий называют молочнокислым брожением: он лежит в основе приготовления кисломолочных продуктов. Анаэробный гликолиз протекает в цитозоле клеток, где содержатся все необходимые для этого ферменты, и не нуждается в митохондриальной дыхательной цепи. АТФ в процессе анаэробного гликолиза образуется за счет реакций субстратного фосфорилирования.
У дрожжей в анаэробных условиях происходит сходный процесс - спиртовое брожение, в этом случае пировиноградная кислота декарбоксилируется с образованием уксусного альдегида, который затем восстанавливается в этиловый спирт:
СН3-СО-СООН → СН3-СНО + СО2;
СН3-СНО + НАД.Н+Н+ → СН3-СН2-ОН + НАД+.
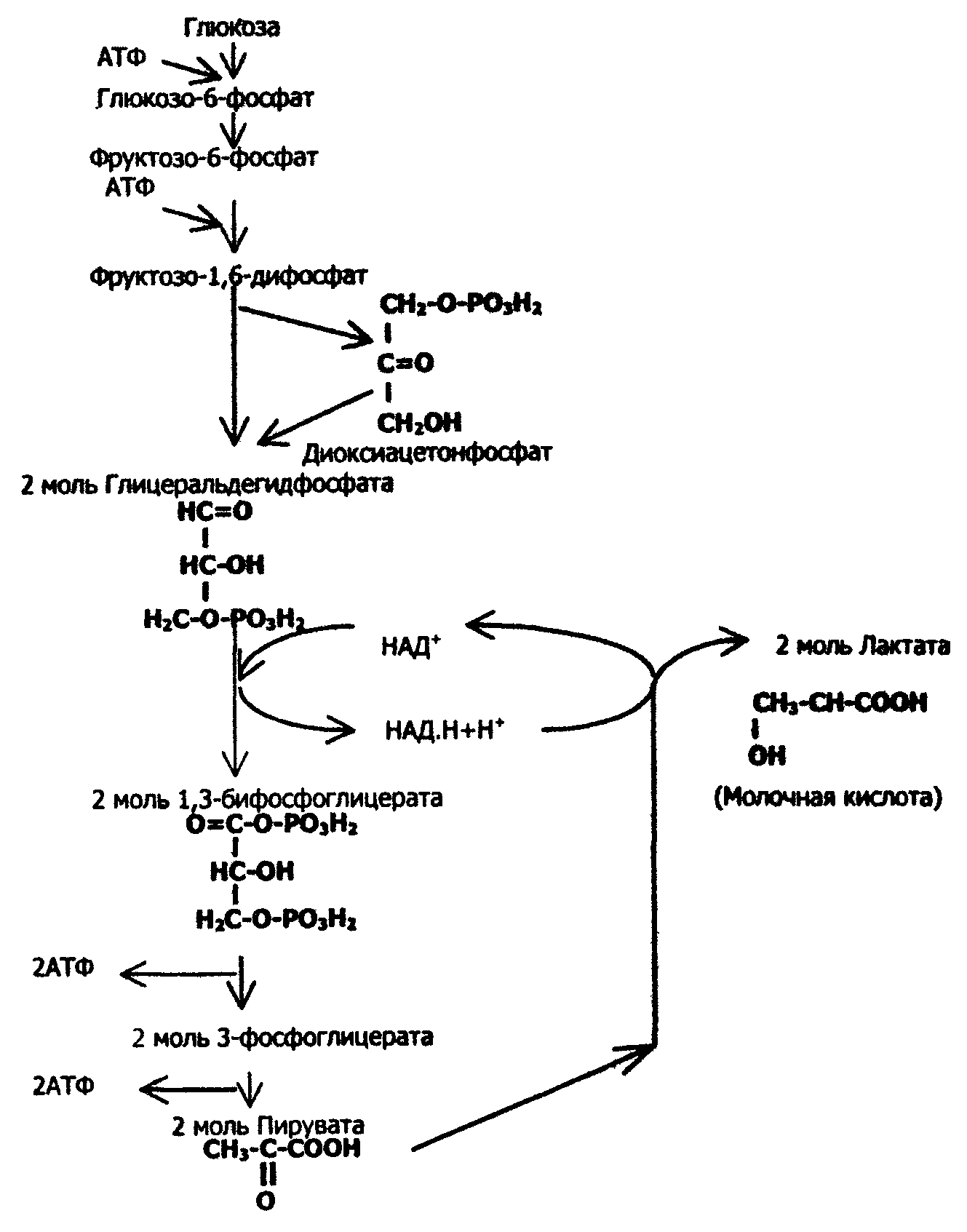
Рис.28. Схема анаэробного гликолиза глюкозы
10.6. Аэробный распад глюкозы
Аэробный распад глюкозы включает в себя три стадии:
1) превращение глюкозы до пировиноградной кислоты (пирувата) - аэробный гликолиз. Эта часть аналогична рассмотренному выше процессу анаэробного гликолиза, за исключением его последней стадии (превращение пирувата в молочную кислоту);
2) общий путь катаболизма;
3) митохондриальная цепь переноса электронов - процесс тканевого дыхания.
Общий путь катаболизма
Общий путь катаболизма сострит из двух этапов.
1-й этап - окислительное декарбоксилирование пировиноградной кислоты. Это сложный многостадийный процесс, катализируемый мультиферментной системой - пируватдегидрогеназным комплексом; локализуется в митохондриях (внутренняя мембрана и матрикс) и может быть выражен суммарной общей схемой:
СН3-СО-СООН + HS-KoA + НАД+ → CH3-CO-SkoA + НАД.Н+Н+ + СО2.
|
пируват |
коэнзим А |
ацетил-КоА |
2-й этап - цикл Кребса (цитратный цикл, или цикл трикарбоновых и дикарбоновых кислот) (рис. 29); локализуется в митохондриях (в матриксе). В этом цикле ацетильный остаток, входящий в ацетил-КоА, образует ряд первичных доноров водорода. Далее водород при участии дегидрогеназ поступает в дыхательную цепь. В результате сопряженного действия цитратного цикла и дыхательной цепи ацетильный остаток окисляется до СО2 и Н2О. Суммарное уравнение всей последовательности превращений глюкозы в ходе аэробного распада следующее:
С6Н12О2 + 6О2 → 6СО2 + 6Н2О
Энергетический эффект аэробного распада - синтез 38 молекул АТФ при расщеплении 1 молекулы глюкозы. Таким образом, в энергетическом отношении полное окисление глюкозы до углекислого газа и воды является более эффективным процессом, чем анаэробный гликолиз. Кислород тормозит анаэробный гликолиз, поэтому в присутствии избытка кислорода наблюдается переход в растительных и животных тканях от анаэробного гликолиза (брожения) к дыханию (аэробный гликолиз), т.е. переключение клеток на более эффективный и экономичный путь получения энергии (эффект Пастера). Роль анаэробного гликолиза в обеспечении организма энергией особенно велика при кратковременной интенсивной работе, когда мощности механизма транспорта кислорода к митохондриям недостаточно для обеспечения аэробного гликолиза. Так, бег в течение ~ 30 секунд (на 200 м) полностью обеспечивается анаэробным гликолизом, при этом скорость анаэробного гликолиза с учащением дыхания уменьшается, а скорость аэробного распада увеличивается. Через 4-5 мин. бега (1,5 км) - половину энергии дает анаэробный, половину аэробный процесс. Через 30 мин. (10 км бега) - энергия поставляется почти целиком аэробным процессом.
Эритроциты вообще не имеют митохондрий, и их потребность в АТФ полностью удовлетворяется за счет анаэробного гликолиза.