

10.2. Метаболизм глюкозы
Транспорт глюкозы из крови в клетки зависит от гормона поджелудочной железы инсулина. При пищеварении концентрация глюкозы в крови повышается (алиментарная гиперглюкоземия), и это стимулирует секрецию гормона в кровь. Инсулин увеличивает проницаемость плазматической мембраны клеток для глюкозы, в результате чего ускоряется перенос глюкозы из крови в клетки. Поступление глюкозы в клетки зависит от инсулина во всех органах, кроме мозга и печени: скорость поступления глюкозы в клетки этих органов определяется ее концентрацией в крови.
Общая схема метаболизма глюкозы представлена на рис. 25.

Рис.25. Схема метаболизма глюкозы
Первым химическим превращением глюкозы в клетках является ее фосфорилирование, в результате взаимодействия с АТФ:
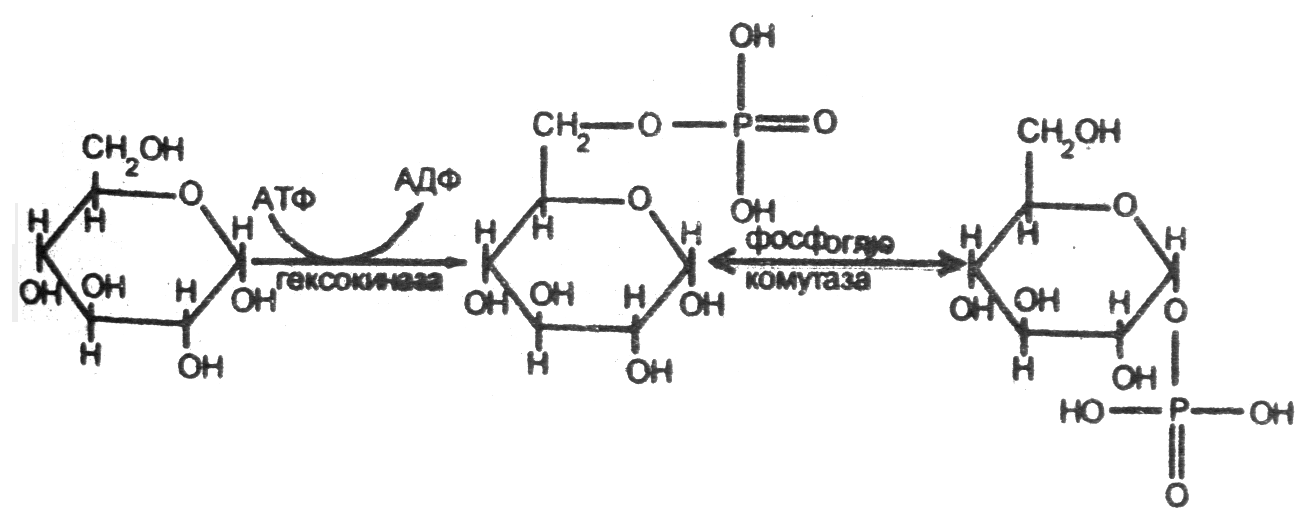
Образующийся глюкозо-6-фосфат не способен проходить через клеточные мембраны - глюкоза как бы «запирается» в клетке в результате фосфорилирования. Возможно и обратное превращение глюкозо-6-фосфата в глюкозу при действии глюказо-6-фосфатазы:
Глюкозо-6-фосфат + H2O → глюкоза + Н3РО4.
Данный фермент есть в печени, в почках, в клетках эпителия кишечника. В других органах и тканях его нет, и, следовательно, проникновение глюкозы в клетки этих органов необратимо. Глюкозо-6-фосфат может обратимо превратиться в глюкозо-1-фосфат при участии фосфоглюкомутазы.
Галактоза и фруктоза, поступающие из кишечного тракта, под действием соответственно галактокиназы и фруктокиназы фосфорилируются по первому углеродному атому:
Галактоза + АТФ → Галактозо-1-фосфат + АДФ;
Фруктоза + АТФ → Фоктозо-1-фосфат + АДФ.
10.3. Биосинтез гликогена
Избыток глюкозы в организме накапливается в виде гликогена, образующегося в процессе гликогенеза практически во всех клетках организма, но больше всего в клетках печени и мышц (в печени - от 2 до 6%, в мышцах - от 0,5 до 2%). Синтез гликогена представляет собой многостадийный процесс (рис.26).
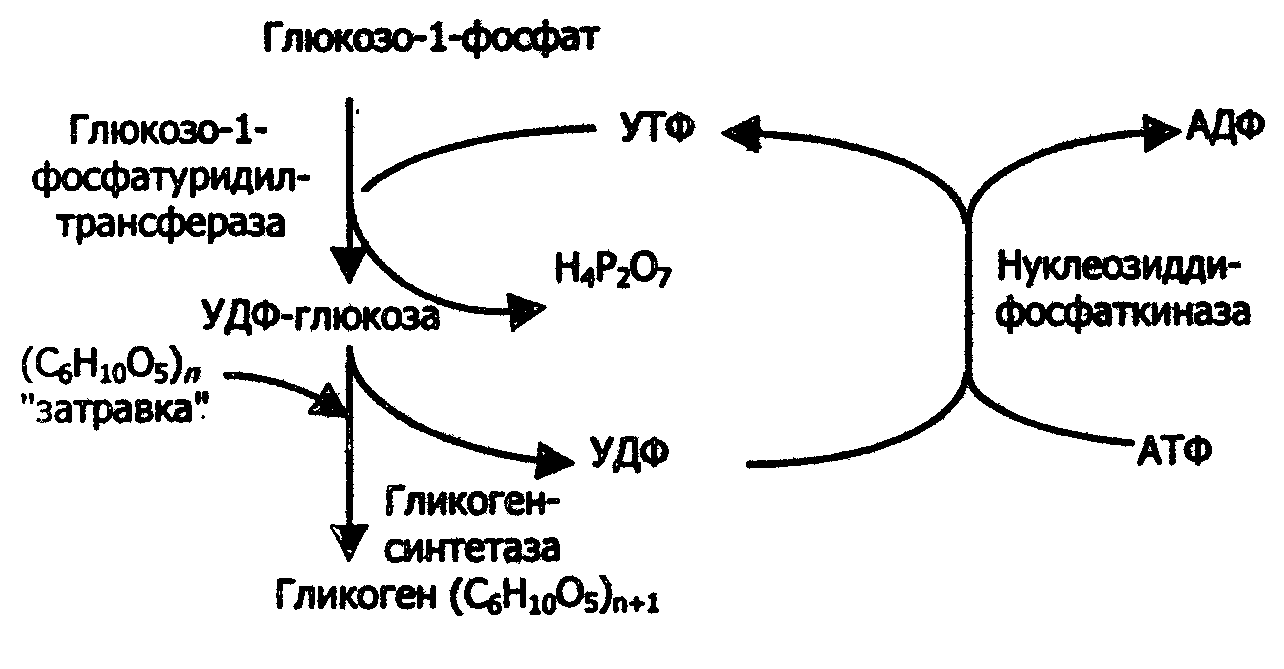
Рис. 26. Схема синтеза гликогена
Донором глюкозных остатков при биосинтезе гликогена служит уридиндифосфатглюкоза (УДФ-глюкоза) – продукт взаимодействия глюкозо-1фосфата и УТФ:

Акцептором глюкозных остатков с УДФ - глюкозы являются олигосахариды из трех или более остатков глюкозы (так называемый "затравочный" глмкоген):
![]()
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и, таким образом, весь цикл превращений глюкозо-1-фосфата начинается сначала.
Глюкоза растворима в воде, а образующийся гликоген образует в воде коллоидный раствор. В клетке он находится не в растворенном состоянии, а в виде гранул диаметром 40-200 нм, включающих одну или несколько молекул.
10.4. Распад гликогена
В процессе гликогенеза клетки забирают глюкозу из крови, а печень восполняет недостаток глюкозы посредством гликогенолиза, т.е. расщепления гликогена:
![]()
Глюкозный остаток отщепляется в виде глюкозо-1-фосфата, последний под действием фосфоглюкомутазы превращается в глюкозо-6-фосфат, дальнейшая судьба которого в печени и в мышцах неодинакова. В печени глюкозо-6-фосфат при участии глюкозо-6-фосфатазы превращается в глюкозу (сущность глюкогенеза), глюкоза выходит в кровь и используется в других органах и тканях. В мышцах нет фермента - фосфатазы, поэтому глюкозо-6-фосфат используется здесь же в мышцах, распадаясь аэробным или анаэробным путем. На рис.27 показаны общие пути синтеза и распада гликогена.
Итак, глюкоза циркулирует в крови и накапливается в печени и мышечных клетках в виде гликогена. Гликоген печени может гидролизоваться для снабжения крови глюкозой, гликоген мышечных тканей до глюкозы не гидролизуется. Можно считать, что сохранение постоянства концентрации глюкозы в крови прежде всего, есть результат одновременного протекания двух процессов: поступление глюкозы в кровь из печени (глюкогенез) и потребление ее из крови тканями, где она используется в основном как энергетический материал.

Рис. 27. Схема метаболизма гликогена:
1-фосфорилаза; 2-фосфоглюкомутаза; З-глюкозо-6-фосфатаза; 4-гексокиназа; 5-глюкозо-1-фосфатуридилтрансфераза; 6-гликогенсинтетаза
В тканях, в том числе и в печени, существует два основных пути распада глюкозы: анаэробный (не требующий присутствия кислорода) и аэробный (для протекания которого необходим кислород).