

Лекция. Нуклеиновые кислоты
.pdfЛЕКЦИЯ |
НУКЛЕИНОВЫЕ КИСЛОТЫ |
Нуклеиновые кислоты-это биополимеры, |
макромолекулы которых состоят из |
мономерных звеньев- нуклеотидов, поэтому |
нуклеиновые кислотыэто полинуклеотиды, |
цепи которых имеют неразветвленное |
строение. |
Молекулярная масса этих макромолекул колеблется от 25 тыс до 1 |
Молекулярная масса этих макромолекул колеблется от 25 тыс до 1 |
млн. |
млн. |
РНК и ДНК – сложные |
высокомолекулярные соединения, но в |
их состав входит небольшое число |
химических компонентов более |
простого состава. |
При полном гидролизе нуклеиновых |
кислот образуются следующие |
химические соединения: |
Нуклеиновые кислоты обеспечивают хранение и |
передачу наследственной информации и |
непосредственно участвуют в механизмах реализации |
этой информации путем программирования синтеза |
всех клеточных белков. |
Мономерной единицей нуклеиновых |
кислот являются нуклеотиды. |
Существуют 2 разновидности |
полинуклеотидов, входящих в состав |
живой клетки: рибонуклеиновая (РНК) и |
дезоксирибонуклеиновая (ДНК) |
кислоты. |
ПРОДУКТЫ ГИДРОЛИЗА |
|
РНК |
ДНК |
H3PO4 |
H3PO4 |
Д-рибоза |
2-дезокси-Д-рибоза |
Аденин |
Аденин |
Гуанин |
Гуанин |
Цитозин |
Цитозин |
Урацил |
Тимин |
1
►Итак, в состав нуклеиновых кислот |
|
|
входят: |
1. |
Углеводный остаток |
2. |
Гетероциклическое основание |
3. |
Фосфатная группа |
Составы РНК и ДНК отличаются: |
||
1. Углеводным компонентом. РНК содержит Д- |
||
рибозу-(-β-Д-рибофуранозу) |
|
|
А в молекуле ДНК углевод представлен 2-дезокси- |
||
Д-рибозой-(-β-2-дезокси-Д-рибофуранозой) |
||
|
HOH2C |
OH |
R=OH -D-рибофураноза |
O |
|
R=H |
2-Дезокси- -D- |
|
рибофураноза |
|
|
|
HO |
R |
2.Гетероциклическим основанием. |
||||
В РНК содержится урацил, а в ДНК- тимин. |
||||
|
O |
|
|
|
|
|
|
|
O |
HN3 4 5 |
HN |
CH3 |
||
2 |
|
6 |
|
|
1 |
|
|
||
O |
N |
|
O |
N |
|
H |
|
|
H |
Урацил Ura |
|
Тимин Thy |
|
|
|
(5-метил-2,4-диоксопиримидин, |
|||
(2,4-диоксопиримидин) |
5-метилурацил) |
|
||
|
Пиримидиновые основания |
|
|||
|
|
|
Пиримидин |
|
|
|
|
|
N |
|
|
|
O |
|
N |
|
NH2 |
|
|
|
|
|
|
HN3 4 5 |
|
O |
|
N |
|
|
CH3 |
|
|
||
2 |
|
HN |
|
|
|
1 6 |
|
O |
N |
||
|
|
||||
O |
N |
O |
N |
|
|
|
|
|
H |
||
|
H |
|
Тимин Thy |
|
|
|
|
H (5-метил-2,4- |
Цитозин Cyt |
||
|
|
|
|||
Урацил Ura |
|
диоксопиримидин, |
(4-амино-2- |
||
(2,4-диоксопиримидин) |
5-метилурацил |
оксопиримидин) |
|||
Азотистые основания, входящие в |
|||||
|
состав РНК И ДНК. |
|
|
||
► Азотистые основания(нуклеиновые)- |
|
|
|
||
гетероциклические соединения, производные пурина и |
|||||
пиримидина. |
|
|
|
|
|
|
Пуриновые основания |
|
NH2 |
|
|
|
|
|
|
|
|
|
|
|
N1 |
6 5 |
N |
|
N |
|
7 |
||
N |
|
2 |
4 |
8 |
|
|
|
|
3 |
9 |
|
|
|
|
N |
||
|
|
|
|
N |
|
|
|
O |
|
АденинH |
|
|
NH |
|
|
||
N |
HN |
N |
(6-аминопурин) |
||
|
|
|
|
||
Пурин |
|
|
N Гуанин Gua |
|
|
|
H2N |
N |
|
||
|
|
|
H (2-амино-6-оксопурин) |
||
Гидроксипроизводные |
гетероциклического ряда способны к |
лактим-лактамной таутометрии. |
Однако, при физиологических условиях |
нуклеиновые основания существуют |
только в лактамной форме(ОКСО- |
форме).Обе таутомерные формы имеют |
плоское строение и сохраняют |
ароматичность. |
Ароматичность- основа относительно |
высокой термодинамической |
стабильности гетероциклов. |
2
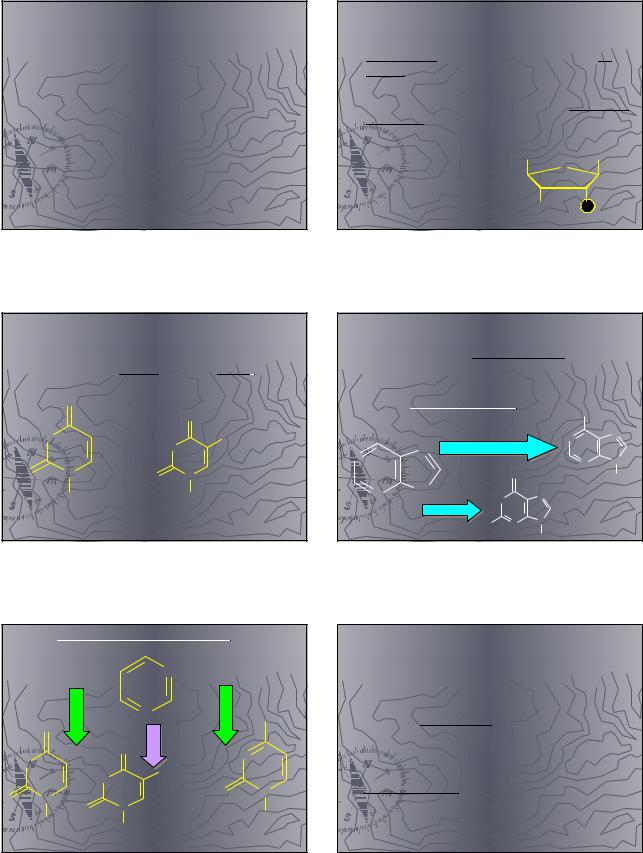
Лактим-лактамная |
таутомерия: |
|
||||||
|
OH |
|
|
O |
|
|
O |
H |
|
|
|
|
|
|
|
|
|
|
N |
N |
H |
N |
N |
H |
|
N |
|
|
N |
||||||
|
|
|
|
|
|
|||
HN |
N |
N |
H2N |
N |
N |
HN2 |
N |
N |
2 |
|
|
|
|
H |
|
||
|
|
H |
|
|
|
|
|
|
Нуклеозиды |
Нуклеозиды сокращенно обозначают |
начальной буквой их латинского |
названия с добавление префикса d в |
случае дезоксинуклеозидов. |
Например, А-аденозин, dА- |
дезоксиаденозин. |
Связь в нуклеозидах осуществляется между |
Связь в нуклеозидах осуществляется между |
С1(аномерным) атомом рибозы(или дезоксирибозы) |
С1(аномерным) атомом рибозы(или дезоксирибозы) |
и атомом азота N1 пиримидинового и N9 |
и атомом азота N1 пиримидинового и N9 |
пуринового оснований. |
пуринового оснований. |
Нуклеозиды |
Нуклеозидыэто N-гликозиды, образованные |
нуклеиновыми основаниями и β-Д- |
рибофуранозой(РНК) или 2-дезокси-β-Д- |
рибофуранозой(ДНК). |
Общая структура нуклеозидов: |
Х = ОН — рибонуклеозиды; |
Х = ОН — рибонуклеозиды; |
Х = Н — дезоксирибонуклеозиды; |
Х = Н — дезоксирибонуклеозиды; |
R - пуриновое или пиримидиновое основание: |
R - пуриновое или пиримидиновое основание: |
Названия нуклеозидов производятся от тривиального |
названия соответствующего гетероциклического |
азотистого основания с суффиксами |
–идину пиримидиновых |
–идину пиримидиновых |
–озин у пуриновых |
–озин у пуриновых |
нуклеозидов. |
нуклеозидов. |
Например: |
Например: |
Цитозин+Рибоза=Цитидин |
Цитозин+Рибоза=Цитидин |
Аденин+Дезоксирибоза=Аденозин |
Аденин+Дезоксирибоза=Аденозин |
(Исключение составляет название ТИМИДИН, а не |
(Исключение составляет название ТИМИДИН, а не |
дезокситимидин!) |
дезокситимидин!) |
ГИДРОЛИЗ НУКЛЕОЗИДОВ |
►Нуклеозиды(N-гликозиды) расщепляются |
в КИСЛОЙ среде,но устойчивы в |
СЛАБОЩЕЛОЧНОЙ. |
U + H2O |
U + H2O |
H+ Урацил+β,Д-рибофураноза |
H+ Урацил+β,Д-рибофураноза |
ОН- |
ОН- |
Реакция не идет |
Реакция не идет |
3

Лекарственные средства нуклеиновой |
природы. |
В клетках в свободном состоянии |
содержатся некоторые нуклеозиды, не |
являющиеся компонентами нуклеиновых |
кислот. Они обладают антибиотической |
активностью и приобретают всё большее |
значение при лечении злокачественных |
образований. Известны несколько десятков |
таких нуклеозидов, выделенных |
микроорганизмов, растительных и |
животных тканей. |
Названия нуклеотидов РНК |
1.Уридин-5’-фосфат (5’-уридиловая кислота) |
1.Уридин-5’-фосфат (5’-уридиловая кислота) |
2.Цитидин-5’-фосфат (5’-цитидиловая кислота) |
2.Цитидин-5’-фосфат (5’-цитидиловая кислота) |
3.Аденозин-5’-фосфат (5’-адениловая кислота) |
3.Аденозин-5’-фосфат (5’-адениловая кислота) |
4.Гуанозин-5’-фосфат (5’-гуаниловая |
4.Гуанозин-5’-фосфат (5’-гуаниловая |
кислота) |
кислота) |
Нуклеотиды имеют |
большое значение не |
только как |
строительный материал |
для нуклеиновых кислот. |
Они участвуют в |
биохимических |
процессах, тесно |
связаны с ферментами |
и необходимы для |
проявления |
ферментативной |
активности. |
Нуклеотиды |
Нуклеотидыэто фосфаты |
нуклеозидов. |
Фосфорная кислота обычно этерифицирует |
спиртовой гидроксил при С-5’ или при C-3’ в |
остатке углевода. |
Любой нуклеотид состоит из 3х компонентов: |
-углевода |
-углевода |
-нуклеинового основания |
-нуклеинового основания |
-остатка фосфорной кислоты |
-остатка фосфорной кислоты |
Для связывания этих компонентов используется |
Для связывания этих компонентов используется |
N-гликозидная и сложноэфирная связь. |
N-гликозидная и сложноэфирная связь. |
Названия нуклеотидов ДНК |
Названия нуклеотидов ДНК |
1.Тимидин-5’-фосфат (5’-тимидиловая кислота) |
1.Тимидин-5’-фосфат (5’-тимидиловая кислота) |
2.Дезоксицитидин-5’-фосфат (5’-дезоксицитидиловая |
2.Дезоксицитидин-5’-фосфат (5’-дезоксицитидиловая |
кислота) |
кислота) |
3.Дезоксиаденозин-5’-фосфат (5’-дезоксиадениловая |
3.Дезоксиаденозин-5’-фосфат (5’-дезоксиадениловая |
кислота) |
кислота) |
4.Дезоксигуанозин-5’-фосфат (5’-дезоксигуаниловая |
4.Дезоксигуанозин-5’-фосфат (5’-дезоксигуаниловая |
кислота) |
кислота) |
Гидролиз нуклеотидов |
При частичном гидролизе нуклеотидов |
отщепляется остаток фосфорной кислоты и |
образуются нуклеозиды. |
4
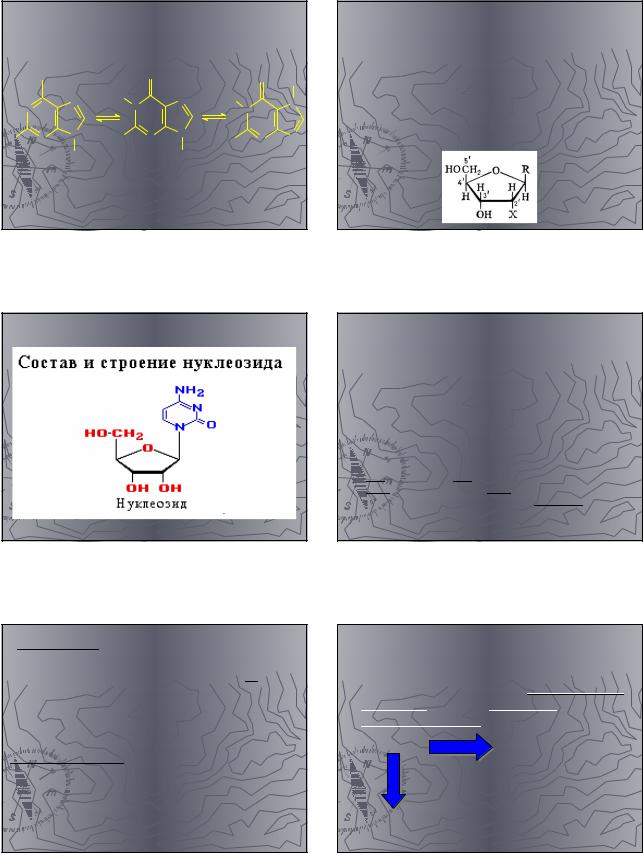
Кроме рассмотренных нуклеотидов |
существует еще 2 типа фосфорных |
эфиров нуклеотидов: |
Фосфорная кислота: этерифицирует |
одновременно две ОН-группы |
углеводного остатка в одном и том же |
нкулеотиде. Они сокращенно называются |
цикломонофосфаты |
Практически во всех клетках |
|
присутствуют два |
|
нуклеозидциклофосфата: |
|
|
сАМФ-обладает высокой |
|
биологической |
|
активностью. |
|
АМФ циклический (cAMP) является |
|
АМФ циклический (cAMP) является |
|
универсальным посредником передачи |
|
универсальным посредником передачи |
|
внутриклеточного сигнала с внешней |
|
внутриклеточного сигнала с внешней |
|
стороны клеточной мембраны к |
|
стороны клеточной мембраны к |
|
эффекторным системам клетки, |
|
эффекторным системам клетки, |
|
действие которых вызывает |
аденозин-3',5'- |
действие которых вызывает |
генерализованную реакцию клетки на |
|
циклофосфат |
генерализованную реакцию клетки на |
воздействие внешнего биологически- |
|
сАМФ(цАМФ) |
воздействие внешнего биологически- |
активного вещества, например |
|
|
активного вещества, например |
|
гормона. |
|
гормона. |
Эти соединениявыполняют важную роль |
коферментов большого числа ферментов |
дегидрогеназ и, следовательно, являются |
участниками окислительно-восстановительных |
реакций. В соответствии с этим они могут |
существовать как в окисленной(НАД+,НАДФ+), |
так и в восстановленной(НАДН,НАДФН) |
формах. |
Фосфорная кислота связывает 2 |
|
нуклеотида в одну молекулу, образуя в |
|
молекулах между собой ангидридную |
|
связь: |
|
| |
| |
- Р - О – Р – |
|
|| |
|| |
O |
O |
Наиболее важным представителем этой группы |
|
является никотинамид- |
|
адениндинуклеотид(НАД) и его фосфат НАДФ. |
|
НАД и НАДФ |
Никотинамидадениндинуклеотидфосфат, |
Никотинамидадениндинуклеотидфосфат, |
НАДФ (рифосфопиридиннуклеотид (ТПН))- широко |
распространённый в природе кофермент.Структура НАДФ |
установлена в 1934 О. Варбургом. |
Подобно никотинамидадениндинуклеотиду обнаружен во |
всех типах клеток; |
Участвует в реакциях окисления — восстановления: |
1. служит акцептором водорода при окислении главным |
образом углеводов; |
2. в восстановленной форме является донором водорода при |
биосинтезе жирных кислот. |
В хлоропластах растительных клеток НАДФ восстанавливается |
при световых реакциях фотосинтеза и затем обеспечивает |
водородом синтез углеводов при темновых реакциях. |
5
НАД и НАДФ |
Окисление, восстановление |
В ходе биологического дегидрирования, |
являющегося особым случаем окисления, |
субстрат теряет 2 атома водорода,т.е. 2 |
протона и 2 электрона(2Н+,2е-)или протон |
и гидридион(Н+ и Н-).Кофермент НАД+ |
обычно рассматривается как акцептор |
гидрид-иона Н-.В результате |
восстановления за счет присоединения |
гидрид-иона НАД+ пиридиниевое кольцо |
переходит в 1,4-дигидропиридиновый |
фрагмент. Данный процесс обратим. |
Нуклеозидполифосфаты |
Во всех тканях организма в свободном |
состоянии содержатся моно-,ди- |
трифосфаты нуклеозидов.Особенно |
широко известны аденинсодержащие |
нуклеотиды- |
аденозин-5-фосфат(АМР),аденозин-5- |
дифосфат(ADP) и аденозин-5- |
трифосфат(АТР).В ряде биохимических |
реакций участвуют также нуклеотиды,как |
гуанозинтрифосфат(GTP),уридинтрифосф |
ат(UTP),цитидинтрифосфат(CTP). |
Нуклеотиды,фосфорилированные в |
разной степени,способны к |
взаимопревращениям путем |
наращивания или отщепления |
фосфатных групп.Дифосфатная |
группа содержит одну,а |
трифосфатная-две ангидридные |
связи,называемые |
макроэргическими,поскольку |
они обладают большим запасом |
энергии.Необходимые для |
образования такой связи |
энергетические затраты |
покрываются за счет |
энергии,выделяющейся в |
процессе метаболизма |
углеводов.При расщеплении |
макроэргической связи |
выделяется~32кДж/моль.С этим |
связана важнейшая роль АТФ |
как «поставщика» энергии во |
всех живых клетках. |
Нуклеозидполифосфаты |
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
NH2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
N |
|
N |
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
HO |
P |
O |
CH2 |
|
|
N |
|
N |
HO |
P |
O |
P |
O |
CH2 |
Ade |
|
O |
|
|
|
|||||||||||||
|
HO |
|
|
|
|
|
|
+H3PO4 |
|
|
|
|
O |
+H3PO4 |
||
|
|
|
|
|
|
|
|
HO |
|
HO |
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-H3PO4 |
||
|
|
|
|
|
|
|
|
|
|
-H3PO4 |
|
|
|
|
|
|
|
|
|
|
OH |
|
|
OH |
|
|
|
Ангидридная |
|
OH |
OH |
||
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
Аденозинмонофосфат |
|
|
|
|
группа |
|
|
|
|||||
|
|
|
|
|
АМФ |
|
|
|
|
|
|
|
|
|
Аденозиндифосфат |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
АДФ |
|
|
|
|
O |
|
O |
|
O |
|
|
|
|
|
|
|
|
|
|
|
HO |
P |
O |
P |
O |
P |
O |
CH2 |
O |
Ade |
|
|
|
|
|
|
|
|
HO |
|
HO |
|
HO |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
Ангидридные группы |
|
OH |
OH |
|
|
|
|
|
|||||
|
|
|
|
|
Аденозинтрифосфат |
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
АТФ |
|
|
|
|
|
|
|
|
|
6

Нуклеозидполифосфаты в |
биохимических процессах |
С участием АТФ и АДФ в организме |
осуществляется важнейший биохимический |
процесс-перенос фосфатных |
групп.Многочисленные реакции |
фосфорилирования можно разделить на 2 |
группы: |
к одной относятся реакции,ведущие к |
образованию сложноэфирной связи; |
к другой-реакции,ведущие к образованию |
ангидридной связи. |
Образование сложных эфиров(фосфатов)- |
|||
типичная реакция в метаболизме |
|||
углеводов.Например,все стадии |
|||
гликолиза(превращение глюкозы в |
|||
пируват)осуществляютя с соединениями только в |
|||
фосфатной форме.Получение фосфатов |
|||
гидроксисодержащих соединений можно |
|||
представить в виде общей схемы: |
|||
|
|
O |
|
ROH + АТФ |
RO |
P |
OH + АДФ |
|
|
OH |
|
Нуклеозидполифосфаты в |
|
||||||
биохимических процессах |
|
||||||
|
|
O |
|
O |
|
O |
|
С |
O |
P |
OH |
C |
O |
P |
OH |
|
|
OH |
|
|
|
OH |
|
Сложный эфир |
|
|
Ангидрид |
|
|||
Перевод карбоксилсодержащих соединений в |
ангидридную форму составляет химическую |
основу активации жирных |
кислот,аминокислот,желчных |
кислот,необходимой для участия их в |
последующих превращениях.При этом в состав |
образующихся ангидридов со стороны АТФ |
может входить либо остаток фосфорной |
кислоты(ацилфосфаты),либо остаток |
АМФ(замещенные ацилфосфаты |
ациладенилаты).Например,при получении |
глутамина,играющего роль в обмене азота |
В организме, |
одной из промежуточных стадий |
является образование |
АЦИЛФОСФАТА. |
Нуклеозидполифосфаты в биохимических процессах. |
|||||||||||
В организме процесс активации карбоксилсодержащих соединени |
|||||||||||
осуществляется также за счет образования ациладенилатов- |
|||||||||||
смешанных ангидридов карбоновых кислот и АТФ(пример |
|||||||||||
взаимодействия галактозы с АТФ): |
|
|
|
|
|
|
|||||
HOCH2 |
|
|
|
|
|
|
|
|
|
|
|
HO |
O |
|
O |
|
O |
|
O |
|
|
|
|
OH |
OH + |
HO |
P |
O |
P |
O |
P |
O |
CH2 |
O |
Ade |
|
|
|
-O |
|
-O |
|
-O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
OH |
|
|
|
|
|
|
|
|
|
|
D-галактоза |
|
|
|
АТФ |
|
|
OH |
|
OH |
||
|
|
|
|
|
|
|
|
||||
HOCH2 |
|
|
|
|
|
|
|
|
|
|
|
HO |
O |
O |
|
|
|
O |
|
O |
|
|
|
|
OH |
OP |
O- |
+ |
-O |
P |
O |
P |
O |
CH2 |
Ade |
|
|
O- |
|
|
|
-O |
|
-O |
|
|
O |
|
|
|
|
|
|
|
|
|
|||
|
OH |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
OH |
OH |
1-фосфат D-галактопиранозы |
|
|
|
|
|
|
АДФ |
|
|
||
|
O |
|
|
|
|
O |
|
O |
|
|
|
|
-OOC CHCH2CH2 |
C |
O- |
+ |
АТФ |
-OOC |
CHCH2CH2 C |
O |
P |
|
O- |
+ АДФ |
|
NH3 |
|
|
|
|
|
NH3 |
|
O- |
|
|
|
|
Глутамат |
|
|
|
|
|
|
|
|
|
|
|
|
|
нуклеофильная атака |
|
|
|
|
|
|
|
|
|||
|
O |
|
O |
|
|
|
O |
|
|
|
O |
|
-OOC CHCH2CH2 |
C |
O |
P |
O- + NH3 |
-OOC CHCH2CH2CNH2 |
+ |
-O |
|
P |
O- |
||
NH3 |
|
|
O- |
|
NH3 |
|
|
|
|
O- |
|
|
|
|
|
|
|
|
Глутамин |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
O |
|
RCOO- + АТФ + KoASH |
|
фермент |
RCO |
SKoA + АМФ + -O |
P |
|
O |
P |
O- |
|||
|
|
|
|
|
|
|
|
O- |
|
|
O- |
|
7
O |
|
O |
|
O |
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
RC |
-O |
P |
O |
P |
O |
P |
O |
CH2 |
O |
|
|
Ade |
|
|
O- |
|
-O |
|
-O |
|
-O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
Ацилат |
|
|
|
АТФ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
OH |
|
|
OH |
|
|
|
O |
|
O |
|
|
|
|
|
|
|
|
O |
|
O |
|
RС |
O |
P |
O |
CH2 |
|
O |
|
Ade |
+ |
-O |
P |
O |
P |
O- |
|
|
-O |
|
|
|
|
|
|
|
O- |
O- |
|
||
|
|
|
|
|
|
|
|
|
|
|
||||
Ациладенилат |
|
|
|
|
|
|
|
|
Дифосфат-ион |
|
||||
|
|
|
|
OH |
|
OH |
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
Примером фундаментального процесса |
|
|
|
||||||||
|
жизнедеятельности,в котором происходит образование |
|
|||||||||||||
|
|
ациладенилатов,служит биосинтез белка.Сначала |
|
|
|||||||||||
|
аминокислота,участвующая в синтезе белка,подвергается |
||||||||||||||
|
O |
|
O |
|
O активации с помощью АТФ. |
|
|
|
|
||||||
-O |
P |
O |
P |
O |
P |
O |
CH2 |
|
Ade |
+ +NH3 CH |
|
COO- |
|
|
|
|
-O |
|
-O |
|
-O |
|
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
R |
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
АТФ |
|
|
|
OH |
OH |
|
-аминокислота |
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
O |
|
|
|
|
|
O |
|
O |
|
|
|
+NH3 |
CH |
C |
O |
P |
O |
CH2 |
|
|
Ade |
|
|
||
|
|
O |
|
P |
O |
P |
O- |
||||||||
|
|
|
R |
O |
|
-O |
|
|
|
+ -O |
|||||
|
|
|
|
|
|
|
|
|
O- |
|
O- |
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
Ангидридная группа |
OH |
|
OH |
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
Аминоациладенилат |
|
|
|
|
|
|
|||
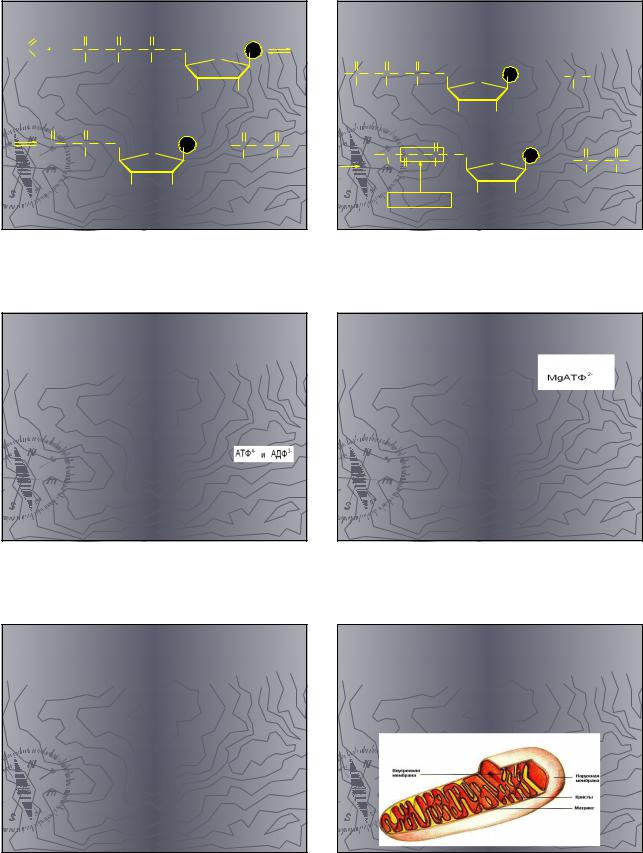
Состояние АДФ и АТФ в |
организме |
Фосфатные группы |
полифосфатов АТФ и АДФ в |
организме находится в |
ионизованном состоянии |
Активная форма АТФ |
Комплексы с магнием |
В ферментативных реакциях |
фосфорилирования различных |
биосубстратов с образованием |
органических фосфатов. |
Концентрация АТФ |
Поддерживается на относительно |
постоянном уровне, его образования |
приблизительно уравновешивается |
скоростью его гидролиза. |
В организме человека около 30 г АТФ. |
Образование АТФ в клетке |
Происходит в митохондриях за счет |
энергии, выделяющейся при |
биологическом окислении. |
8

Нуклеиновые кислоты |
Френсис Крик и Джеймс Уитсон рядом со своей моделью ДНК |
(© A. Barrington Brown/Science Source/Photo Researchers, Inc.) |
Активация нуклеиновых кислот |
Происходит при образовании |
лабильных комплексов с |
внутриклеточными катионами |
СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ |
|||||||||||
|
|
|
|
NH2 |
|
|
|
|
|
|
5'-Конец |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
N |
|
|
N |
|
|
|
|
|
|
|
|
|
|
|
|
A |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
пентоза основание |
|
O CH2 |
|
N |
|
N |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
O |
|
|
|
|
|
|
|
|
|
фосфат |
|
|
|
|
|
NH2 |
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|||
O |
|
|
|
|
|
N |
|
C |
|
|
пентоза основание |
|
|
|
|
|
|
|
|
|
|
|
|
O P |
O |
CH2 |
|
|
N |
|
O |
|
|
|
фосфат |
O |
|
|
|
|
O |
|
|||||
OH |
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
N |
|
|
пентоза основание |
|
|
|
|
|
|
|
|
|
|
NH |
||
|
|
|
|
|
|
|
|
|
|
||
|
|
O |
|
|
|
|
|
|
|
|
G |
|
|
|
|
|
|
|
N |
|
|
фосфат |
|
|
|
O P |
O |
CH2 |
|
|
N |
NH2 |
|||
|
|
O |
|
|
|
|
|
||||
|
|
OH |
|
|
|
|
|
|
O |
пентоза основание |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
H3C |
NH |
фосфат |
|
|
|
|
|
O |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
T |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
P |
O |
CH2 |
O |
N |
O |
|
|
|
|
|
|
OH |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3'-Конец |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
Первичная структура |
|
|
|
|
|
|
|
O |
P |
|
|
|
d(…A—С—G—Т...) |
|
|
|
участка цепи ДНК |
|||||||
|
|
OH |
|
|
|||||||
СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ |
ДНК бактериофага Т2 |
ДНК была |
высвобождена из |
головки фага с |
помощью |
осмотического шока. |
Слева— |
микрофотография |
целой частицы фага. |
Снимки 1962 года |
ПЕРВИЧНАЯ СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ |
► Под первичной структурой нуклеиновых кислот понимают порядок, |
последовательность расположения мононуклеотидов в полинуклеотидной цепи |
ДНК и РНК. Такая цепь стабилизируется 3',5'-фосфодиэфирными связями. |
Поскольку молекулярная масса нуклеиновых кислот колеблется в широких |
пределах (от 2•104 до 1010–1011), установить первичную структуру всех |
известных РНК и особенно ДНК весьма сложно. Тем не менее во всех |
нуклеиновых кислотах (точнее, в одноцепочечной нуклеиновой кислоте) |
имеется один и тот же тип связи – 3',5'-фосфодиэфирная связь между |
соседними нуклеотидами. |
► Установлено, что в образовании межнуклеотидной связи участвуют |
гидроксильные группы в 3'- и 5'-положениях остатков углевода. Эту |
общую основу структуры можно представить следующим образом: |
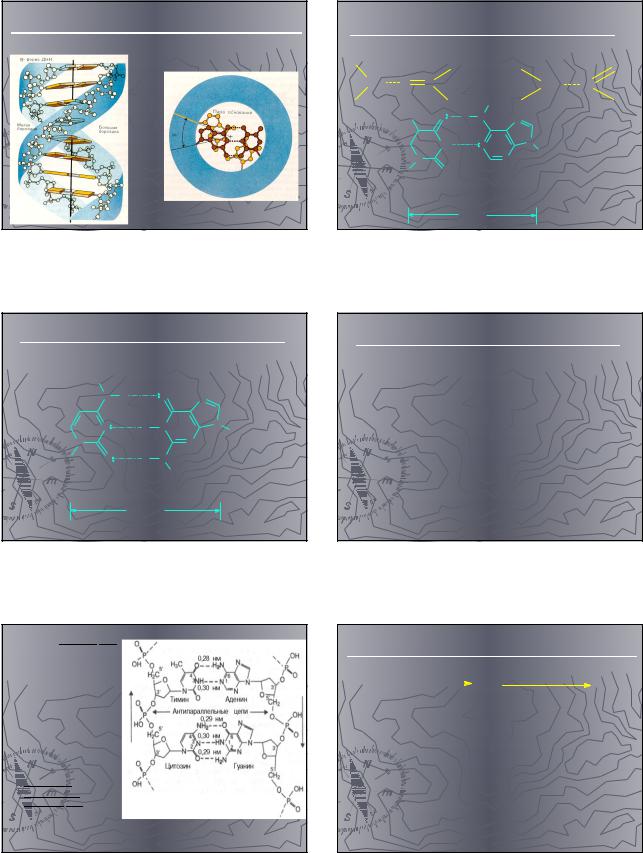
Вторичная структура |
нуклеиновых кислот |
► В соответствии с моделью Дж. |
Уотсона и Ф. Крика, |
предложенной в 1953 г. на |
основании ряда аналитических |
данных, а также |
рентгеноструктурного анализа |
молекула ДНК состоит из двух |
цепей, образуя правовращающую |
спираль, в которую обе |
полинуклеотидные цепи |
закручены вокруг одной и той же |
оси. Удерживаются цепи |
благодаря водородным связям, |
образующимся между их |
азотистыми основаниями (рис. |
3.1). Обе цепи поли-нуклеотидов |
в биспиральной молекуле ДНК |
имеют строго определенное |
пространственное расположение, |
при котором азотистые основания |
находятся внутри, а |
фосфорильные и углеводные |
компоненты – снаружи. |
9
СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ |
Вторичная структура нуклеиновых кислот |
Вторичная структура нуклеиновых кислот |
|||||
|
H |
|
|
|
|
|
N |
H |
|
O |
N |
|
N |
|
H |
N |
N |
|
|
К цепи |
|||
|
|
|
|
|
|
|
N |
|
|
|
N |
К цепи |
O |
|
H |
N |
|
|
|
|
|
|
H |
|
Цитозин |
|
|
|
Гуанин |
|
|
|
1,08 |
нм |
|
Обе цепи в молекуле ДНК |
имеют |
противоположную |
полярность. Это |
означает, что |
межнуклеотидная связь |
в одной цепи имеет |
направление 5'–>3', а в |
другой – 3'–>5'. |
Подобная |
направленность цепей |
имеет важное |
биологическое |
значение при |
репликации и |
транскрипции |
молекулы ДНК. |
Вторичная структура нуклеиновых кислот |
|||||
NH |
O |
C |
|
NH |
N |
|
|
|
|
H |
|
|
H3C |
O |
H N |
N |
|
|
|
NH |
N |
N |
|
|
|
К цепи |
|
||
|
|
N |
|
|
|
|
|
|
N |
|
|
|
К цепи |
O |
|
|
|
|
|
Тимин |
|
Аденин |
|
|
|
|
1,11 нм |
|
|
Вторичная структура нуклеиновых кислот |
Правила Чаргаффа |
1) количество пуриновых оснований равно |
1) количество пуриновых оснований равно |
количеству пиримидиновых оснований; |
количеству пиримидиновых оснований; |
2) количество аденина равно количеству тимина; |
2) количество аденина равно количеству тимина; |
количество гуанина равно количеству цитозина; |
количество гуанина равно количеству цитозина; |
3) количество оснований, содержащих аминогруппу |
3) количество оснований, содержащих аминогруппу |
в положениях 4 пиримидинового и 6 пуринового |
в положениях 4 пиримидинового и 6 пуринового |
ядер, равно количеству оснований, содержащих в |
ядер, равно количеству оснований, содержащих в |
этих же положениях оксогруппу. Это означает, что |
этих же положениях оксогруппу. Это означает, что |
сумма аденина и цитозина равна сумме гуанина и |
сумма аденина и цитозина равна сумме гуанина и |
тимина. |
тимина. |
БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕИНОВЫХ КИСЛОТ |
|
ДНК Транскрипция РНК Трансляция |
Белок |
1. Информационная, или матричная |
|
РНК (ее обозначают мРНК) считывает и |
|
переносит генетическую информацию |
|
от ДНК, содержащейся в хромосомах, к |
|
рибосомам, где происходит синтез |
|
белка со строго определенной |
|
последовательностью аминокислот. |
|
10