

Вопрос 3. Влияние паразита на популяцию хозяев
Успех существования биотрофного паразита должен зависеть от того, насколько эффективно он конкурирует за ресурсы с теми частями организма хозяина, которые им поражены. Такая внутренняя конкуренция обусловливает, по крайней мере отчасти, снижение выживаемости, плодовитости, роста «внешней» конкурентоспособности хозяина. Влияние степени поражения паразитами на смертность хозяев показано на рис. 6, а у зараженных клещом Hydraphantes tenuabilis клопов-водомерок Hydrometra myrae снижается выживаемость и замедляются размножение и развитие (рис. 7). Менее заметное влияние паразитизма выражается в том, что зараженные хозяева становятся более уязвимыми для хищников. Например, бакланы избирательно добывают тех особей плотвы, которые зараженных ленточным червем Ligula intestinalis.
Во всех случаях наносимый вред отражает взаимодействие между самим заражением хозяина паразитами и другим фактором или факторами. В случае с плотвой - инвазия и нападение хищника; заболевания людей усугублялись недостатком пищи: детская смертность в слаборазвитых странах, по-видимому, в основном связана с болезнями, которые при хорошем питании в развитых странах не смертельны. При этом никогда не следует забывать о роли, которую играет в них генетическая изменчивость. Некоторые болезни, в частности, корь, лишь недавно проникли в Африку и Индию. Они поражают население, предки которого никогда не подвергались отбору на устойчивость или невосприимчивость к этим инфекциям, тем более - в условиях плохой обеспеченности пищей.
Паразиты и популяционная динамика хозяев
В принципе, в отношении взаимодействий между паразитами и хозяевами могут быть сделаны примерно те же выводы, что и при исследовании отношений хищник-жертва и фитофаг-растение. Паразиты вредят особям хозяина с интенсивностью, зависящей от плотности обеих популяций. Кроме того, зараженные и незараженные особи хозяина проявляют компенсаторные реакции, которые могут существенно снижать поражение их популяции в целом. Теоретически предсказуемы несколько исходов взаимодействия: варьирующая степень сокращения плотности популяции хозяина, варьирующая степень устойчивости и амплитуды колебаний численности и даже множественных состояний равновесия.

Рис. 6. Влияние паразитов на состояние популяций хозяина.

Рис. 7. Зависимости показателей жизнеспособности клопа-водомерки от степени зараженности паразитом.
При исследовании системы паразит-хозяин возникает ряд трудностей: (1) паразиты часто вызывают ухудшение «здоровья» своего хозяина, не приводя к его немедленной гибели, поэтому обычно трудно отделить их влияние от действия другого фактора или факторов, с которыми возбудитель взаимодействует; (2) даже если паразит является причиной гибели хозяина, без детального обследования трупа это нередко трудно установить (особенно в случае микропаразитов); (3) кроме того, паразитологи, раньше стремились изучать биологию того или иного паразита, не обращая особенного внимания на то, как он влияет на популяцию хозяина в целом, а экологи не были склонны заниматься паразитами. С другой стороны, фитопатологи, медицинские и ветеринарные паразитологи по вполне понятным причинам выбирали для изучения наиболее опасных паразитов, обычно связанных с плотными и агрегированными популяциями хозяев, не слишком интересуясь гораздо более распространенными взаимоотношениями паразитов с популяциями видов, менее зависимых от человека. Возможно, самый определенный вывод, касающийся роли паразитов в популяционной динамике хозяев, - это то, что оценка ее является одной из важнейших проблем, стоящих перед экологией.
Безусловно, в некоторых случаях снижение плотности популяции хозяев под влиянием паразитов хорошо выражено. Широко распространенное и интенсивное использование опрыскиваний, прививок и лекарств в растениеводстве и ветеринарии - все это свидетельство ущерба, ожидаемого в отсутствии борьбы с заболеваниями (в медицине основное внимание часто направлено не на увеличение численности популяции, а на сохранение отдельных особей). Снижение плотности популяции хозяев под влиянием паразитов можно наблюдать и в контролируемых лабораторных условиях (рис. 8). Для естественных популяций такие непосредственно сравнимые результаты получить необычайно трудно. Даже если одна популяция заражена, а другая нет, незараженная популяция наверняка существует в иных условиях и, не исключено, что сама она заражена другим паразитом, который в первом случае отсутствует или встречается редко.

Рис. 8. Результаты лабораторных экспериментов по подавлению роста численности популяций хозяина паразитами.
Модели, оценивающие степень снижения плотности популяции хозяина, были разработаны Андерсоном (Anderson, 1979) (рис. 9). В случае скоротечных заболеваний, когда зараженные хозяева не выздоравливают (например, для многих микропаразитов растений и беспозвоночных), наибольшее снижение плотности хозяев происходит при низких-умеренных уровнях патогенности (рис. 9,А и Б). Высокая патогенность ведет к тому, что многие хозяева гибнут, не передав возбудителя, который в итоге исчезает, и популяция хозяина достигает предельной для данной среды плотности. С другой стороны, при появлении патогенности и возрастания ее даже до очень невысокого уровня, плотность популяции хозяев быстро падает. Кроме того, плотность хозяев в большей степени ограничена их репродуктивными возможностями (рис. 9,А), чем высокой скоростью заражения паразитами (рис. 9,Б). Сходная картина (рис. 9,В) возникает и в случае устойчивых инфекций (например, большинства макропаразитарных). Хорошо видно, что по мере увеличения агрегированности паразитов, их влияние на плотность популяции хозяев уменьшается.
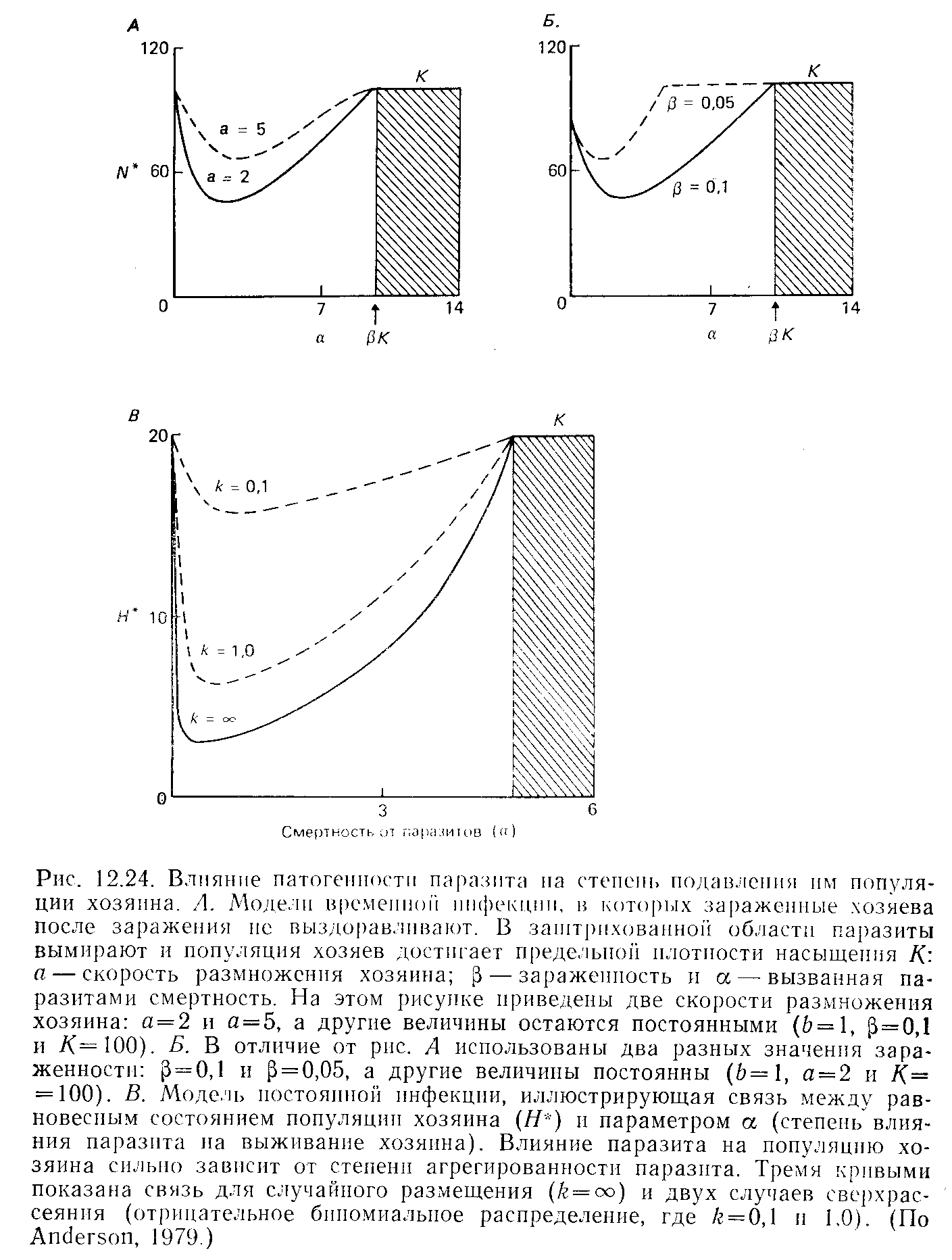
Рис. 9. Три модели, описывающие влияние паразита на хозяина при различных величинах патогенности.
Итак, в целом паразиты должны снижать плотность популяций своих хозяев, причем сильнее всего при низкой и умеренной патогенности. Этот результат заслуживает внимания с точки зрения все более частого использования паразитов для борьбы с вредителями.
Наконец, растет число данных, свидетельствующих о том, что болезнями можно объяснить (по крайней мере частично) сложные картины популяционной динамики хозяина, особенно циклические колебания. Например, на рис. 10 показаны такие колебания, наблюдавшиеся в альпийской популяции вредителя лиственницы Zeiraphera ditiiana на фоне изменений частоты зараженности вирусом гранулеза. Аналогичное совпадение можно получить, например, в модели взаимодействия лиса-вирус бешенства, описывающей трех-пятилетние циклические колебания численности млекопитающего даже в отсутствие более обычных естественных циклов, а также для популяции шотландской куропатки, зараженной нематодой Trichosirongyltis tenuis на севере Англии.

Рис. 10. Сопоставление динамики численности лиственничной моли и встречаемости паразитических грануловирусов.