

лекция
.docЛекция
Первичные фотопроцессы в биологических системах Исследования молекулярных механизмов фотопревращений родопсина и бактериородопсина представляют собой важную область биофизики фотобиологических процессов. Бактериородопсин был обнаружен в пурпурной мембране галофильных бактерий, которая способна преобразовывать энергию света. Эта система, видимо, является наиболее простой из всех изученных систем запасания световой энергии в форме разности электрохимических потенциалов Н+ . Зрительный пигмент родопсин, так же как и бактериородопсин, — практически единственный белок в фоторецепторной мембране зрительной клетки сетчатки глаза (на его долю приходится до 80% всего белка в мембране). По целому ряду свойств родопсин и бактериородопсин схожи (аминокислотный состав, молекулярная масса, химическая природа хромофора). Общие черты присущи и процессам фотохимических превращений, особенно на первых стадиях.
Фотопревращения бактериородопсина
Как уже было сказано, бактериородопсин обнаружен в пурпурной мембране галобактерий. Мембранный комплекс с бактериородопсином способен запасать световую энергию в форме разности электрохимических потенциалов.
Бактериородопсин содержит один хромофор-ретиналь (полиеновый альдегид) в комплексе с белком – опсином. Опсин использует энергию света для активного переноса протонов через мембрану, в результате происходит синтез АТФ и выполнение других функций. Ретиналь ковалентно связан с опсином, образуя так называемое шиффовооснование, которое может быть протонировано.
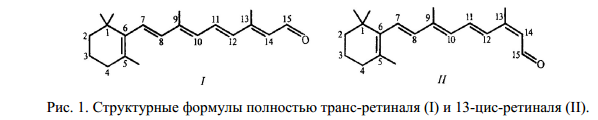
После поглощения кванта света ретиналь изомеризуется и переходит из полностью транс- в 13-цис форму (рис. 1). Протон с шиффово основания переносится на аминокислотный остаток, вакансия заполняется протоном с другого остатка. По эстафетной цепочке транспортных реакций протон освобождается на внешней стороне мембраны, а поглощается из внеклеточного пространства.
Конформационные переходы бактериородопсина инициируются кулоновским взаимодействием в донорно-акцепторных парах аминокислот. После поглощения кванта конформационная энергия запасается в виде градиента электрохимического потенциала ионов водорода.
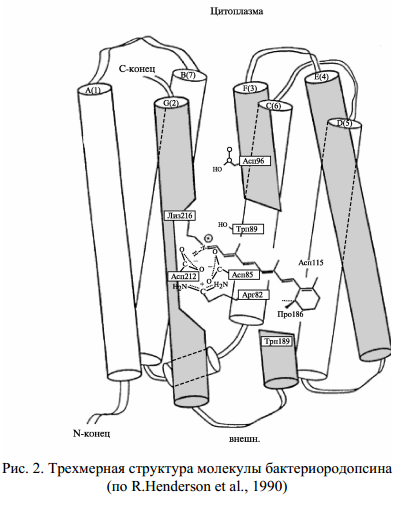
Трансмембранная часть бактериородопсина сложена из 7 регулярных α-спиралей, идущих от одного до другого края мембраны, а одинокая β-шпилька и все нерегулярные участки цепи (соединяющие спирали-петли) выходят из мембраны. Сидящие на α-спиралях гидрофобные группы обращены «наружу» к липидам (тоже гидрофобным) мембраны. Полярные же группы (их немного) обращены внутрь очень узкого канала, по которому идет протон. Ретиналь прикреплен к лиз-216, находящемуся в конце Gα-спирального столба (рис. 2). В состав ближайшего белкового окружения ретиналя помимо лиз-216 входят и другие аминокислоты, которые играют важную роль в структурной стабилизации и функционировании бактериородопсина. Они могут играть роль сложного противоиона для протона шиффова основания.
Фотоцикл бактериородопсина
В основе функционирования бактериородопсина как протонной помпы лежит его способность за счет энергии света переносить протоны с цитоплазматической на внешнюю сторону клеточной мембраны. После поглощения кванта света ретиналь изомеризуется и переходит из полностью-транс в 13-цис форму (рис..1). При этом протон с шиффова основания переносится на Асп 85, после чего образовавшаяся вакансия заполняется протоном, перешедшим с Асп 96. Такой внутримолекулярный перенос протона инициирует эстафетную цепочку транспортных реакций, в результате которых происходит высвобождение протона на внешней стороне мембраны и поглощение протона из внутриклеточного пространства. В схематическом виде это показано на рис. 3. В ходе цикла фотохимических превращений бактериородопсин испытывает конформационные переходы, которые по сути сходны с таковыми у большинства ферментов. Эти конформационные переходы инициируются кулоновским взаимодействием в донорно-акцепторных парах аминокислот: между положительно заряженным внутримолекулярным донором и отрицательно заряженным акцептором протона.
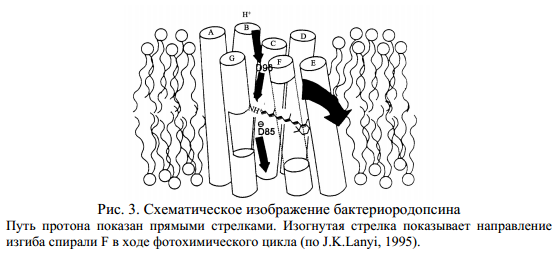
До поглощения кванта света структура белка стабилизирована энергией связи комплекса, образованного протонированным шиффовым основанием, заряженными группами в локусе ретиналя и связанной водой (рис. 2). В этом конформационном состоянии (конформация Е) протон шиффова основания координационно связан с Асп 85 и находится в контакте с наружной стороной мембраны. Фотоизомеризация ретиналя смещает шиффово основание относительно Асп 85, но возникающие в молекуле напряжения способствуют переносу протона с шиффова основания на Асп 85. Бактериородопсин при этом переходит в новую конформацию (конформация С), отличающуюся сдвигом спирали G от цитоплазматической стороны мембраны к центру молекулы (см. рис. 3). В конформации G шиффово основание уже находится в контакте с цитоплазматической стороной мембраны, а его контакт с Асп 85 и, следовательно, с наружной стороной мембраны нарушен.
Таким образом, бактериородопсин представляет собой ионный насос с переменным доступом активного центра к противоположным сторонам мембраны. В терминах конформационной динамики бактериородопсин представляет собой систему с двумя устойчивыми состояниями, причем избыточная конформационная энергия запасается в ходе фотоцикла в виде трансмембранного градиента электрохимического потенциала ионов водорода.
Механизмы переноса протона.
Современные модели переноса протона в цикле бактериородопсина основаны на представлениях о двух протонных каналах, связывающих ретиналь с внутренней и наружной сторонами мембраны. Ретиналь служит своего рода молекулярным переключателем, регулирующим общий трансмембранный перенос протонов по цитоплазматическому и периплазматическому каналам (рис. 3). Происходящая в результате транс-цис фотоизомеризация ретиналя является триггером, «запускающим» всю дальнейшую цепь последовательных превращений фотоцикла. Первичным акцептором протона от шиффова основания является Асп 85. Группа X, определенная как Глю-204, непосредственно отдает протон в наружную среду. Остаток Асп 96 репротонирует шиффово основание, восполняя дефицит протонов за счет цитоплазматического источника.
В результате изменения конфигурации ретиналя (фотоизомеризация) шиффово основание смещается в гидрофобную область белка с более низким значением диэлектрической постоянной. Это приводит к увеличению взаимодействия между положительным протоном основания и отрицательным зарядом противоиона Асп 85, в результате чего сродство к протону шиффова основания уменьшается. Теперь протон может перейти на отрицательно заряженный остаток Асп 85 и протонировать его. Протонирование Асп 85 и нейтрализация отрицательного заряда его группы СОО~понижает сродство к протону аминогруппы, что способствует ее депротонированию и выбросу протона во внешнюю среду.
Основы зрительной рецепции позвоночных
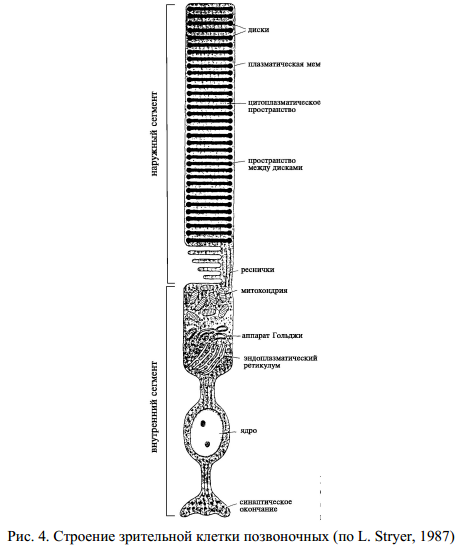
Светочувствительные клетки сетчатки – палочки и колбочки генерируют зрительный сигнал в ответ на поглощение квантов света (рис. 4). Основной молекулярной фоточувствительной структурой зрительной клетки является фоторецептурная мембрана.
В фосфолипидном бислое мембраны встроены молекулы зрительного пигмента родопсина. В темноте ретиналь-хромофор родопсина находится в 11-цис-положении. Поглощение кванта света переводит его в транс-положение (рис. 5).
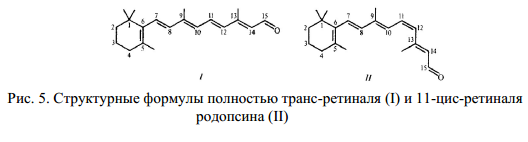
Под действием света резко уменьшается проницаемость мембраны для ионов натрия, один квант закрывает 100-300 каналов. Возникающий светоиндуцированный электрический сигнал гиперполяризации передается к синаптическому окончанию на внутреннем сигменте палочки и приводит к появлению нервного импульса. Изомеризация ретиналя приводит к закрытию натриевых каналов при участии большого числа молекул медиатора (посредника), находящихся в цитоплазме. Натриевые каналы способен открывать цГМФ (3, 5 – циклический гуанозин-монофосфат). Между возбуждением родопсина и гидролизом цГМФ фосфодиэстеразой существует сложная многоступенчатая связь.
Фотохимические превращения родопсина. Родопсин – хромопротеин с молекулярной массой порядка 40000. Большой гидрофобный сегмент находится внутри мембраны, гидрофильный сегмент – на поверхности мембраны.
Хромофором является ретиналь, представляющий собой половину молекулы β-каротина. Ковалентная связь ретиналя с опсином осуществляется при образовании альдиминной связи, которая представляет собой протонированное шиффово основание, находящееся в гидрофобном окружении:
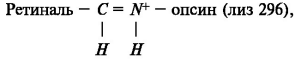
В исходном состоянии в родопсине ретиналь находится в 11-цис, 12-в-цис-конфигурации. В результате фотолиза происходит цис-транс-изомеризация хромофора и депротонирование шиффова основания. Эти процессы сопровождаются изменением взаимодействия ретиналя с опсином и конформации последнего. Таким образом, депротонирование шиффова основания служит механизмом его электростатического включения. Основным фактором здесь является нейтрализация заряда боковой цепи Глю 113. Ретиналь, переходя в состояние полностью-транс, запускает этот процесс. На конечной стадии разрывается альдиминная связь и высвобождается свободный ретиналь в транс-конфигурации.