Включение других углеводов в процесс гликолиза фруктоза.
Установлено, что фруктоза, присутствующая в свободном виде во многих фруктах и образующаяся в тонком кишечнике из сахарозы, всасываясь в тканях, может подвергаться фосфорилированию во фруктозо-6-фосфат при участии фермента гексокиназы и АТФ.
АТФ АДФ

 Фруктоза
Фруктозо-6-фосфат
Фруктоза
Фруктозо-6-фосфат
Эта реакция ингибируется глюкозой. Образовавшийся фруктозо-6-фосфат либо превращается в глюкозу через стадии образования глюкозо-6-фосфата и последующего отщепления фосфорной кислоты, либо подвергается дальнейшим превращениям. Из фруктозо-6-фосфата под влиянием 6-фосфофруктокиназы и АТФ образуется фруктозо-1,6-бифосфат.
 АТФ
АДФ
АТФ
АДФ
Ф руктозо-6-фосфат
Фруктозо-1,6-бифосфат
руктозо-6-фосфат
Фруктозо-1,6-бифосфат
Фосфофруктокиназа
Далее фруктозо-1,6-бифосфат может подвергаться дальнейшим
превращениям по пути гликолиза. Таков главный путь включения фруктозы
в мышечной ткани, почках, жировой ткани.
В печени, однако, для этого существует другой путь. В печени имеется
фермент фруктокиназа, который катализирует фосфорилирование фруктозы
не по 6-му, а по 1 -му атому углерода.
 АТФ
АДФ
АТФ
АДФ
 Фруктоза
Фруктозо-1 -фосфат
Фруктоза
Фруктозо-1 -фосфат
Фруктокиназа
Эта реакция не блокируется глюкозой. Образовавшийся фруктозо-1-фосфат расщепляется затем под действием кетозо-1-фосфатальдолазы на диоксиацетонфосфат и В-глицеральдегид.
Образовавшийся О-глицеральдегид под влиянием соответствующей киназы (триоксикиназы) подвергается фосфорилированию до глицеральдегид-3-фосфата. В этот же промежуточный продукт гликолиза переходит и диоксиацетонфосфат.
Существует врожденная аномалия обмена фруктозы, или эссенциальная фруктозурия, которая связана с врожденным недостатком фермента фруктокиназы, т.е. в организме не образуется фруктозо-1 -фосфат. В результате обмен фруктозы возможен только путем фосфорилирования до фруктозо-6-фосфата, но эта реакция тормозится глюкозой, вследствие чего фруктоза накапливается в крови. Почечный порог для фруктозы очень низок, поэтому фруктозурия обнаруживается уже при концентрации фруктозы в крови, равной 0,73 ммоль/л.
Рис. 5 Метаболизм фруктозы
1 - гексокиназа
2 - фосфофруктокиназа
3 - фруктозобифосфатальдолаза
4 - фруктокиназа
5 - кетозо- 1-фосфатальдолаза
6 - триокиназа
7 - глюкозофосфатизомераза
8 - глюкозо-6-фосфатаза
9 - триозофосфатизомераза

Галактоза.
Основным источником галактозы является лактоза пищи, которая в пищеварительном тракте расщепляется до галактозы и глюкозы. Обмен галактозы начинается с превращения ее в галактозо-1-фосфат. Эта реакция катализируется галактокиназой с участием АТФ:
АТФ АДФ

 Галактоза
Галактозо-1-фосфат
Галактоза
Галактозо-1-фосфат
галактокиназа
В следующей реакции в присутствии УДФ-глюкозы фермент гексозо-1-фосфат-уридилтрансфераза катализирует превращение галактозо-1-фосфата в глюкозо-1-фосфат, одновременно образуется уридиндифосфатгалактоза (УДФ-галактоза)
Образовавшийся глюкозо-1-фосфат в дальнейшем либо переходит в глюкозо-6-фосфат и далее подвергается уже известным превращениям, либо под влиянием фосфатазы образует свободную глюкозу, а УДФгалактоза подвергается весьма своеобразной эпимеризации:
 УДФ-галактоза
УДф-глюкоза
УДФ-галактоза
УДф-глюкоза
УДФ-глюкозо-
4-эпимераза
Затем УДф-глюкоза-пирофосфорилаза катализирует расщепление УДФ-глюкозы с образованием глюкозо-1-фосфата. О дальнейшей судьбе образовавшегося глюкозо-1-фосфата было сказано выше.
Среди патологических состояний, возникающих в результате нарушения обмена углеводов, важное место занимает галактоземия - рецессивно наследуемое заболевание. При этом заболевании общее содержание моносахаридов в крови повышается главным образом за счет галактозы, достигает 11,1 - 16,6 ммоль/л. Концентрация глюкозы в крови существенно не изменяется. Кроме галактозы, в крови накапливается также галактозо-1-фосфат. Галактоземия приводит к умственной отсталости и катаракте хрусталика. Возникновение данного заболевания у новорожденных связано с недостатком фермента гексозо-1-фосфат-уридилтрансферазы. С возрастом наблюдается ослабление этого специфического нарушения обмена углеводов.
Рис. 6 Метаболизм галактозы.

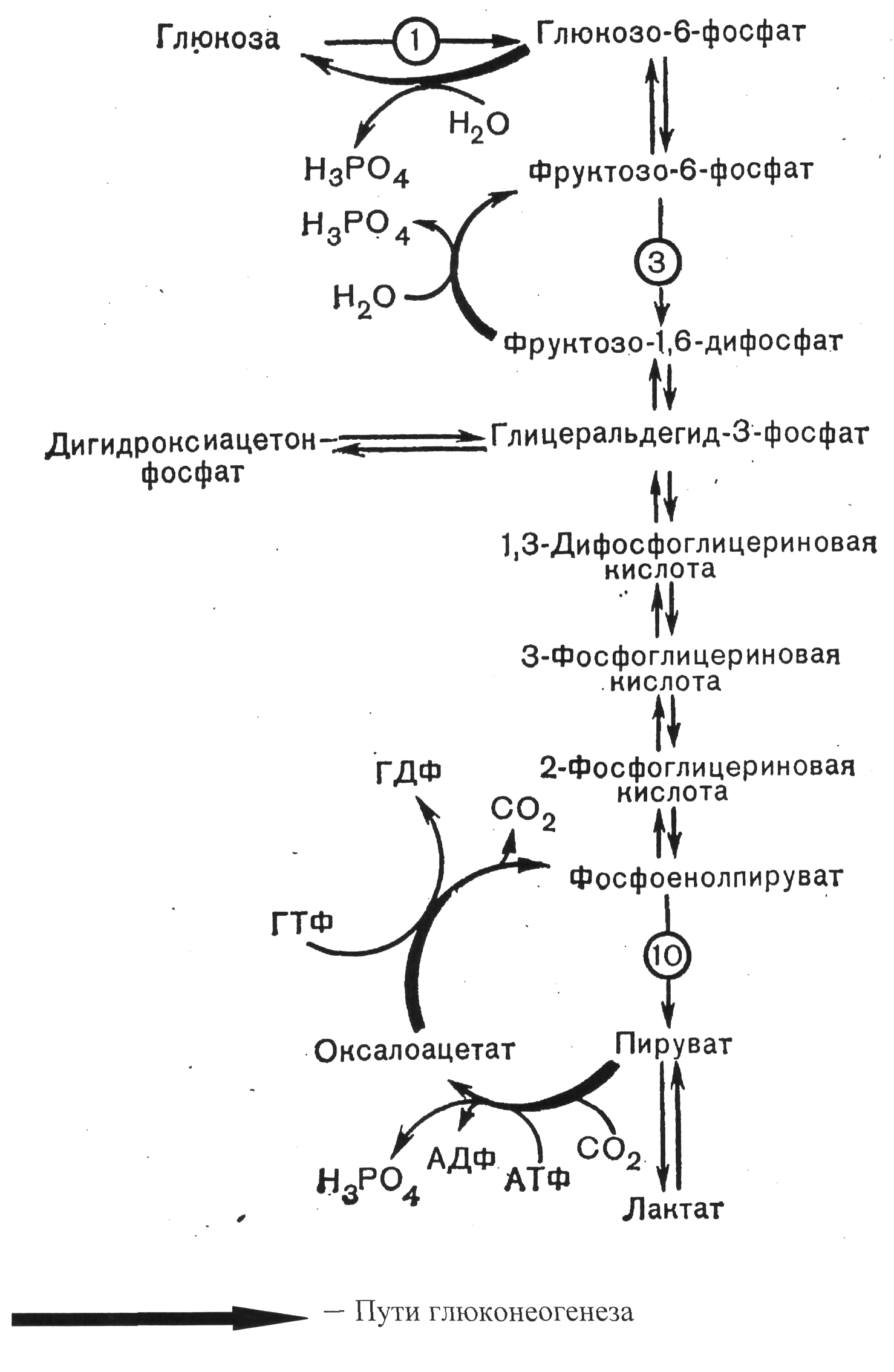
Глюконеогенез.
Глюконеогенез - синтез глюкозы из неуглеводных продуктов. Такими продуктами или метаболитами являются в первую очередь молочная и пировиноградная кислоты, гликогенные аминокислоты, глицерол и ряд других соединений. Иными словами, предшественниками глюкозы в глюконеогенезе могут быть пируват или любое соединение, превращающееся в процессе катаболизма в пируват или один из промежуточных продуктов цикла трикарбоновых кислот. У позвоночных наиболее интенсивно глюконеогенез протекает в клетках печени и почек (корковое вещество).
Большинство стадий глюконеогенеза представляет собой обращение реакций гликолиза. Только три реакции гликолиза (гексокиназная, фосфофруктокиназная и пируваткиназная) необратимы, поэтому в процессе глюконеогенеза на трех этапах используются другие ферменты. Рассмотрим путь синтеза глюкозы из пирувата.
Образование фосфоенолпирувата из пирувата. Синтез фосфоенолпирувата осуществляется в несколько этапов. Первоначально пируват под влиянием пируват-карбоксилазы и при участии С02 и АТФ карбоксилируется с образованием оксалоацетата.
C
 H3
COOH
H3
COOH
C

 =O
+ CO2 + АТФ CH2
+
АДФ
+ H3PO4
=O
+ CO2 + АТФ CH2
+
АДФ
+ H3PO4
C OOHПируваткарбоксилаза
C=O
OOHПируваткарбоксилаза
C=O
COOH
Пируват Оксалоацетат
Затем оксалоацетат в результате декарбоксилирования и фосфорилирования под влиянием фермента фосфоенолпируват-карбоксилазы превращается в фосфоенолпируват. Донором фосфатного остатка в реакции служит гуанозинтрифосфат (ГТФ):
C
 OOH
CH2
OOH
CH2
C


 H2
CHO
PO3H2
H2
CHO
PO3H2
C =O
Фосфоенолпируват-
COOH
=O
Фосфоенолпируват-
COOH
COOH карбоксикиназа
Оксалоацетат Фосфоенолпируват
В дальнейшем было, что в процессе образования фосфоенолпирувата участвуют ферменты цитоплазмы и митохондрий.
Первый этап локализуется в митохондриях. Пируваткарбоксилаза, которая катализирует эту реакцию, является аллостерическим митохондриальным ферментом. В качестве аллостерического активатора данного фермента необходим ацетил-КоА. Мембрана митохондрий непроницаема для образовавшегося оксалоацетата. Последний здесь же в митохондриях восстанавливается в малат.
Реакция протекает при участии митохондриальной НАД-зависимой малатдегидрогеназы. В митохондриях отношение НАДН2/НАД+ относительно велико, в связи с чем внутримитохондриальныи оксалоацетат
легко восстанавливается в малат, который легко выходит из митохондрии, проходя митохондриальную мембрану. В цитоплазме отношение НАДН2/НАД+ очень мало, и малат вновь окисляется в оксалоацетат при участии цитоплазматической НАД-зависимой малатдегидрогеназы.
Дальнейшее превращение оксалоацетата в фосфоенолпируват происходит в цитоплазме клетки.
Рис. 7 Образование фосфоенолпирувата из пирувата.

Превращение фруктозо-1,6-бифосфата во фруктозо-6-фосфат. Фосфоенолпируват, образовавшийся из пирувата, в результате ряда обратимых реакций гликолиза превращается во фруктозо-1,6-бифосфат. Далее следует фосфофруктокиназная реакция, которая необратима. Глюконеогенез идет в обход этой эндергонической реакции. Превращение фруктозо-1,6-бифосфата во фруктозо-6-фосфат катализируется специфической фосфатазой:
Ф руктозо-1,6-бифосфат
+H2O
Фруктозо-6-фосфат + Фн
руктозо-1,6-бифосфат
+H2O
Фруктозо-6-фосфат + Фн
Образование глюкозы из глюкозо-6-фосфата. В последующей обратимой стадии биосинтеза глюкозы фруктозо-6-фосфат превращается в глюкозо-6-фосфат. Последний может дефосфорилироваться (т.е. реакция идет в обход гексокиназной реакции) под влиянием фермента глюкозо-6-фосфатазы:

Глюкозо-6-фосфат + Н2О Глюкоза + Фн
Рис. 8 Общая схема биосинтеза глюкозы из лактата.