

Лекция_5_БИ_М_2014
.pdfГипотеза молекулярных часов.
В 1960е годы - набор данных о последовательностях ряда некрупных белков, находящихся в большом количестве в клетках. (глобины, цитохром с и т.п.)
Для некоторых была показана быстрая скорость накопления замен, а для некоторых – медленная.
Photo from
123 people & Library of Congress
Лайнус Полинг и Эммануэль Марголаш (L.Pauling, E. Margoliash) и коллеги предложили гипотезу молекулярных часов:
Для каждого выбранного белка скорость накопления замен близка к постоянной во всех эволюционных ветвях (родословных).
**исключения – снижение давления отбора в одной из ветвей, интенсивный движущий отбор.
Гипотеза молекулярных часов.
Применение:
Если последовательности белков накапливают замены с постоянными скоростями, то они могут быть использованы для выявления времени дивергенции видов (и последовательностей).
Это аналогично радиоизотопному методу датирования.

Теория нейтральной эволюции.
Распространённое представление об эволюции заключается в том, что так же как организмы, так и молекулы ДНК и белков подвержены естественному отбору.
Согласно теории нейтральной эволюции Мото Кимура
(Motoo Kimura 1968), подавляющее большинство замен нуклеотидов в ДНК не подвержены естественному отбору Дарвиновской эволюции.
Основной причиной эволюционных изменений в последовательностях является
случайный дрейф нейтральных (или почти нейтральных) аллелей.
Естественный отбор присутствует, но имеет для накопления замен ограниченное влияние.
накопление замен при дивергенции C пептида инсулина соответствует нейтральной эволюции.
Неактивные транспозоны и интроны обычно накапливают замены с «нейтральной» скоростью.
Часть генома, которая накапливает замены со скоростью, превосходящей нейтральную обладает исключительной важностью для понимания того, чем например мы отличаемся от мышей
du-cote-de-chez-elysia-chlorotica.blogspot.com
Что такое филогенетическое дерево?
•Филогения - раздел биологии, изучающий родственные взаимоотношения разных групп живых организмов. Филогению отображается обычно в виде "эволюционных древ" или систематических названий.
•Филогенетика (=молекулярная филогенетика) – те же взаимоотношения, но на уровне отдельных белковых (генных) семейств

Молекулярная филогения: Цели.
Молекулярная филогения отвечает на вопросы:
•Сколько генов имеют родство с моим целевым геном?
•Была ли вымершая (в 1880х) Квага больше похожа на лошадь, чем на зебру?
•Был ли Дарвин прав что люди эволюционно ближайшие родственники шимпанзе и горилл?
•В каком родстве состоят киты, дельфины и коровы?
!
Квага
•Когда и где появился ВИЧ?
•Какова история жизни на земле?
Молекулярная филогения: возможности.
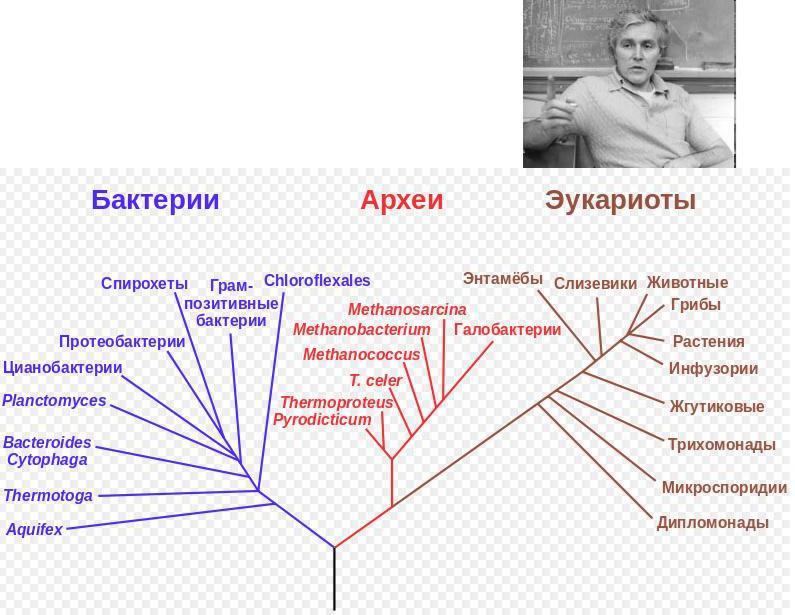
Карл Вёзе (Carl Woese 1970е)
Анализ молекулярной эволюции 5S рРНК. Доказательство существования общего предка ВСЕХ известных организмов.
Последний универсальный общий предок
(англ. last universal common ancestor, LUCA)

Типы данных ?
Molecular (DNA, RNA, proteins)
Morphological (soft tissue, hard tissue, extant, extinct)
Преимущества молекулярных данных
•Наследуемость.
•Недвусмысленность в описании молекулярных характеристик
•Поддаются количественному анализу
•Оценка гомологии легче, чем морфологические исследования
•Данных много
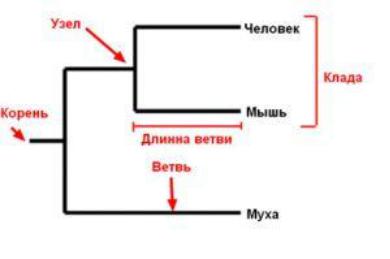
Основные термины
Узел (node) — точка разделения предковой последовательности (вида, популяции) на две независимо эволюционирующие. Соответствует внутренней вершине графа, изображающего эволюцию.
Лист (leaf, OTU – оперативная таксономическая единица) — реальный (современный) объект; внешняя вершина графа. Ветвь (branch) — связь между узлами или между узлом и листом; ребро графа.
Корень (root) — гипотетический общий предок.
Клада (clade) - группа двух или более таксонов или последовательностей ДНК, которая включает как своего общего предка, так и всех его потомков.
Номенклатура филогенетических деревьев.