

Структура рибонуклеиновых кислот (рнк)
Первичная структура РНК – порядок чередования рибонуклеозидмонофосфатов в полинуклеотидной цепи. В РНК, как и в ДНК, нуклеотиды связаны между собой 3',5'-фосфодиэфирными связями. Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная ОН-группа 5'-углеродного атома, на другом конце – ОН-группа 3'-углеродного атома рибозы, поэтому концы называют 5'- и 3'-концами цепи РНК.
Вторичная структура РНК
Молекула рибонуклеиновой кислоты построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли – «шпильки», за счёт водородных связей между комплементарными азотистыми основаниями A-U и G-C. Участки цепи РНК в таких спиральных структурах антипараллельны, но не всегда полностью комплементарны, в них встречаются неспаренные нуклеотидные остатки или даже одноцепочечные петли, не вписывающиеся в двойную спираль. Наличие спирализованных участков характерно для всех типов РНК.
Третичная структура РНК
Одноцепочечные РНК характеризуются компактной и упорядоченной третичной структурой, возникающей путём взаимодействия спирализованных элементов вторичной структуры. Так, возможно образование дополнительных водородных связей между нуклеотидными остатками, достаточно удалёнными друг от друга, или связей между ОН-группами остатков рибозы и основаниями. Третичная структура РНК стабилизирована ионами двухвалентных металлов, например ионами Mg2+, связывающимися не только с фосфатными группами, но и с основаниями.
Основные типы рнк
В цитоплазме клеток присутствуют 3 типа рибонуклеиновых кислот – транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомальные РНК (рРНК). Они различаются по первичной структуре, молекулярной массе, конформации, продолжительности жизни и, самое главное, по функциональной активности.
http://www.biochemistry.ru/biohimija_severina/B5873Part25-141.html
Методы определения первичной и вторичной структуры нуклеиновых кислот
Секвенирование – это общее название методов, которые позволяют установить последовательность нуклеотидов в молекуле ДНК. В настоящее время нет ни одного метода секвенирования, который бы работал для молекулы ДНК целиком; все они устроены так: сначала готовится большое число небольших участков ДНК (клонируется молекула ДНК многократно и «разрезается» её в случайных местах), а потом читается каждый участок по отдельности.
Клонирование происходит либо просто выращиванием клеток в чашке Петри, либо (в случаях, когда это было бы слишком медленно или по каким-то причинам не получилось бы) при помощи так называемой полимеразной цепной реакции. В кратком и неточном изложении работает она примерно так: сначала ДНК денатурируют, т.е. разрушают водородные связи, получая отдельные нити. Затем к ДНК присоединяют так называемые праймеры; это короткие участки ДНК, к которым может присоединиться ДНК-полимераза – соединение, которое, собственно, и занимается копированием (репликацией) нити ДНК. На следующем этапе полимераза копирует ДНК, после чего процесс можно повторять: после новой денатурации отдельных нитей будет уже вдвое больше, на третьем цикле – вчетверо, и так далее.
Все эти эффекты достигаются в основном с помощью изменений температуры смеси из ДНК, праймеров и полимеразы; для наших целей важно, что это достаточно точный процесс, и ошибки в нём редки, а на выходе получается большое число копий участков одной и той же ДНК. Разные методы секвенирования отличаются друг от друга не методами клонирования, а тем, как потом прочесть получившийся «суп» из многочисленных копий одной и той же ДНК.
Метод ДНК-ДНК гибридизации основан на том факте, что стабильность ДНК-ДНК дуплексов при определенной температуре зависит от числа нуклеотидов образующих комплементарные пары. Очевидно, что число комплементарных нуклеотидов в дуплексе где обе нити происходят из одной и той же молекулы ДНК (т.е. в гомодуплексах) равно 100%. Если же обе нити имеют разное происхождение (гетеродуплекс), то, в зависимости от числа произошедших мутаций, число комплементарных пар будет меньше 100%. Соответсвенно гетеродуплексы должны распадаться (плавится) при более низкой температуре, чем гомодуплексы. Причем, чем ниже температура плавления, тем больше различия в двух последовательностях. Температурная стабильность гибридной ДНК определяется температурой при которой 50% гибридной ДНК диссоциировалось в одноцепочечную форму. Затем эта температура сравнивается со средней температурой 50%-го плавления гомодуплексов обоих типов последовательностей участвующих в образовании гетеродуплекса, эта температура обычно обозначается Tm. Разница между медианной температурой плавления гетеро- и гомодуплексов обозначается как dTm. Показана линейная зависимость dTm от числа неспаренных оснований ( Britten et. al., 1974 ): p=cdTm. Константа c обычно определяется условиями проведения эксперимента и обычно варьирует от 0.01 до 0.015. Определение dTm требует большого числа повторений, т.к. велика экспериментальная ошибка.
Основным свойством ДНК является ее способность к репликации.
http://postnauka.ru/longreads/468
1.9. Репликация ДНК, транскрипция, трансляция, обратная транскрипция. Амплификация ДНК. Биосинтез белка, аминокислотный код. Организация генов, строение генов у про- и эукариот, понятие о клонировании.
Репликация – это процесс самоудвоения молекул ДНК, происходящий под контролем ферментов. Репликация осуществляется перед каждым делением ядра. Начинается она с того, что спираль ДНК временно раскручивается под действием фермента ДНК-полимеразы. На каждой из цепей, образовавшихся после разрыва водородных связей, по принципу комплементарности синтезируется дочерняя цепь ДНК. Материалом для синтеза служат свободные нуклеотиды, которые есть в ядре.
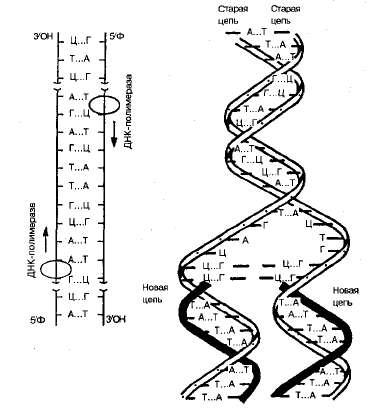
Схема репликации ДНК
Таким образом, каждая полинуклеотидная цепь выполняет роль матрицы для новой комплементарной цепи (поэтому процесс удвоения молекул ДНК относится к реакциям матричного синтеза). В результате получается две молекулы ДНК, у каждой из которых одна цепь остается от родительской молекулы (половина), а другая – вновь синтезированная. Причем одна новая цепь синтезируются сплошной, а вторая – сначала в виде коротких фрагментов, которые затем сшиваются в длинную цепь специальным ферментом – ДНК-лигазой. В результате репликации две новые молекулы ДНК представляют собой точную копию исходной молекулы.
Биологический смысл репликации заключается в точной передаче наследственной информации от материнской клетки к дочерним, что и происходит при делении соматических клеток.
http://sbio.info/page.php?id=11
Литература:
1) Н. Грин, У. Стаут, Д. Тейлор – Биология.
2) З.А. Шабарова и А.А. богданов – Химия нуклеиновых кислот и их полимеров.
3) А.П. Пехов – Биология и общая гинетика.
4) А. Микельсон – Химия нуклеозидов и нуклеотидов.
5) З. Гауптман, Ю. Грефе, Х. Ремане – Органическая химия
Транскри́пция – это процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5'- к 3'- концу, то есть по матричной цепи ДНК РНК-полимераза движется в направлении 3'- 5'.
Транскрипция состоит из стадий инициации, элонгации и терминации. Единицей транскрипции является транскриптон, фрагмент молекулы ДНК, состоящий из промотора, транскрибируемой части и терминатора.
Инициация транскрипции – это сложный процесс, зависящий от последовательности ДНК вблизи транскрибируемой последовательности (а у эукариот также и от более далеких участков генома — энхансеров и сайленсеров) и от наличия или отсутствия различных белковых факторов.
Элонгация транскрипции
Момент перехода РНК-полимеразы от инициации транскрипции к элонгации точно не определен. Три основных биохимических события характеризуют этот переход в случае РНК-полимеразы кишечной палочки: отделение сигма-фактора, первая транслокация молекулы фермента вдоль матрицы и сильная стабилизация транскрипционного комплекса, который кроме РНК-полимеразы включает растущую цепь РНК и транскрибируемую ДНК. Эти же явления характерны и для РНК-полимераз эукариот. Переход от инициации к элонгации сопровождается разрывом связей между ферментом, промотором, факторами инициации транскрипции, а в ряде случаев – переходом РНК-полимеразы в состояние компетентности в отношении элонгации. Фаза элонгации заканчивается после освобождения растущего транскрипта и диссоциации фермента от матрицы (терминация).
На стадии элонгации в ДНК расплетено примерно 18 пар нуклеотидов. Примерно 12 нуклеотидов матричной нити ДНК образует гибридную спираль с растущим концом цепи РНК. По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание, а позади – восстановление двойной спирали ДНК. Одновременно освобождается очередное звено растущей цепи РНК из комплекса с матрицей и РНК-полимеразой. Эти перемещения должны сопровождаться относительным вращением РНК-полимеразы и ДНК.