Генетические карты
В состав генома человека входят две различающиеся по организации и особенностям наследования системы: ядерный геном и митохондриальный геном. В ядре соматических клеток человека содержится около 30 000 пар генов, которые локализованны в 46 молекулах ДНК.
Каждая хромосома содержит в среднем 2000 генов. Гены расположены в линейном порядке и разделены некодирующими последовательностями (сателлитная ДНК, спейсеры). Гены одной хромосомы наследуются совместно. Это явление называется сцеплением генов. Каждая хромосома, таким образом, представляет одну группу сцепления. Митохондриальный геном представлен кольцевыми молекулами ДНК и содержит 37 генов, расположенными очень компактно и наследуемыми по материнской линии.

Явление сцепление характерно только для генов одной и той же хромосомы, в то время как расположенные в разных хромосомах гены наследуются независимо, по законам Менделя. Сцепление бывает полным и неполным. Причина неполного сцепления - кроссинговер, происходящий в мейозе. В ходе кроссинговера происходит взаимный обмен аллельными генами.
Частота кроссинговера неодинакова для различных локусов и может варьировать от 0% до 50%, коррелируя с расстоянием между генами. Чем далее друг от друга расположены гены, тем больше частота кроссинговера между ними и, наоборот. Данное положение позволяет определить расстояние между генами по частоте кроссинговера и лежит в основе составления генетических карт. Эти карты представляют собой графическое изображение хромосом и расположенных на них генов с указанием расстояния между ними.

В настоящее время, благодаря методам дифференциальной окраски и соматической гибридизации, разработаны физические карты хромосом, в которых показано точное расположение генов с указанием расстояния между ними в парах нуклеотидов.
Установление сцепления между генами и определение групп сцепления представляет значительный интерес в медицинской генетике. Примерами сцепленных генов у человека являются: ген фактора Rh и ген эллиптоцитоза; ген АВО и ген пигментной ксеродермы (ХР); гены Duffy и врожденной катаракты; ген MNSs и dentinogenesis imperfecta-1 (DI-1); ген Xg и гены гемофилии А (НЕМА), гемофилии В (НЕМВ), дальтонизмаI (Dalt).

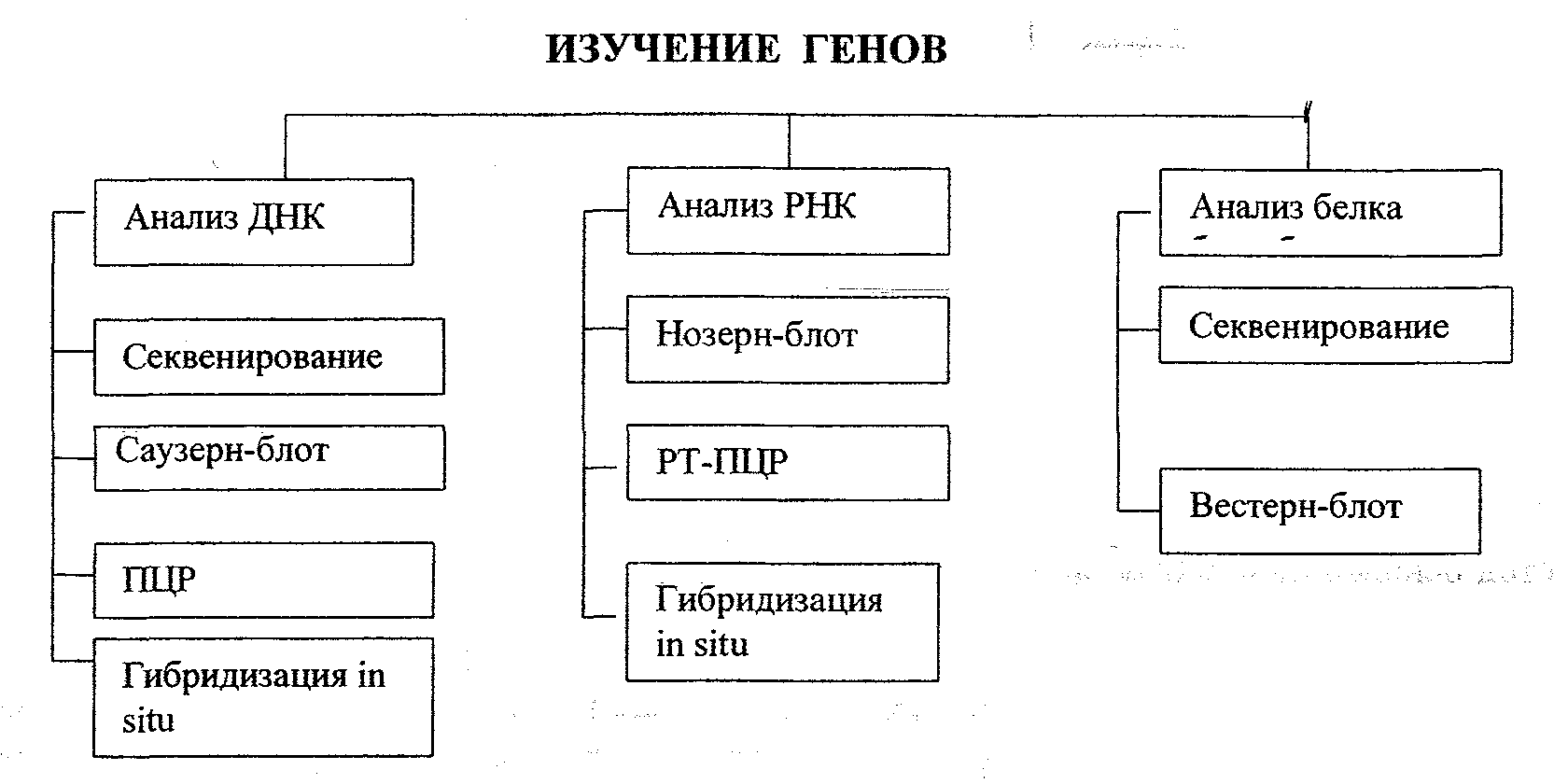
Методы анализа генов
Методы рекомбинантной ДНК создали предпосылку для разработки новых методов молекулярной диагностики с большей разрешающей способностью и, следовательно, более точных и информативных. Об абсолютном преимуществе молекулярного подхода говорит тот факт, что в отличие от других методов диагностики, ограниченных выявлением исключительно фенотипических аспектов, анализ ДНК, направленный непосредственно на изучение генотипа, является единственным методом, изучающим первичные нарушения (мутации), то есть именно первопричину возникновения болезней.
Технология рекомбинантной ДНК позволяет определить нормальные гены и/или мутантные их варианты, установить носителей мутантного гена, диагностировать наследственную патологию до рождения ребенка или в предсимптоматической стадии, а в ближайшем будущем проводить и генную терапию.
Молекулярное изучение генов может осуществляться многими путями в зависимости от поставленной цели:
1. секвестрование ДНК для определения первичной структуры гена;
2. метод Саузерн-блот для определения позиции гена в геноме;
3. метод Нозерн-блот для определения экспрессии генов (анализ мРНК);
4. метод Вестерн-блот для определения белкового продукта гена;
5. метод ПЦР для специфичной амплификации фрагментов ДНК.
В лабораториях молекулярной биологии используются различные варианты перечисленных методов.
Все перечисленные методы основаны на различных принципах манипулирования нуклеиновыми кислотами:
- специфическая рестрикция ДНК с целью получения интересующего фрагмента;
- идентификация фрагментов ДНК или РНК е помощью специальных зондов, комплементарных искомому участку;
- идентификация нормальных и патологических генов путем ПЦР - специфическая реакция отбора праймеров, комплементарных гену/последовательности;
- визуализация необходимого фрагмента по результатам электрофореза и специфического маркирования ДНК/РНК с использованием компьютерных программ чтения и анализа результатов;
- комплексная интерпретация результатов в зависимости от использованного метода.
|
Для разделения полученных фрагментов ДНК применяется электрофорез макромолекул в агарозном или полиакриламидном геле. Будучи отрицательно заряжены, фрагменты молекул нуклеиновых кислот в электрическом поле перемещаются с разной скоростью в зависимости от молекулярной массы. Более короткие фрагменты перемещаются быстрее, в то время, как более длинные - медленнее. Для определения размеров фрагментов в геле, одновременно с интересующими фрагментами помещаются на соседние дорожки и фрагменты - маркеры длины. Молекулы нуклеиновых кислот могут быть обнаружены в геле при окраске флуоресцентным красителем или по радиоактивной метке. В случае радиоактивного мечения, фрагменты определяются с помощью авторадиографии, которая заключается в накладывании на гель светочувствительной пленки. |