

Биохимический этап – цикл Кальвина. С3 путь (Цикл Кальвина)
Цикл состоит из трех этапов:
карбоксилирования,
восстановления,
регенерации первичного акцептора СО2 и синтеза конечного продукта фотосинтеза.
1. Карбоксилирование. Фосфорибулокиназа фосфорилирует при участии АТФ рибулозо-5-фосфат с образованием АДФ и рибулозо-1,5-дифосфата. Последний является акцептором СО2 и под действием рибулозобифосфаткарбоксилазы (РуБФК) присоединяет СО2. В результате образуются 2 молекулы 3-фосфоглицериновой кислоты (3-ФГК). ФГК является первичным продуктом ассимиляции углерода, так как в ее молекуле содержится 3 атома углерода, то этот цикл получил второе название С3.
2. Восстановление. Фосфоглицераткиназа при участии АТФ фосфорилирует 3-ФГК и образовавшаяся 1,3-дифосфоглицериновая кислота восстанавливается с помощью НАДФН и дегидрогеназы фосфоглицеринового альдегида до 3-фосфоглицеринового альдегида (3-ФГА).
3. Регенерация. После фиксации трех молекул СО2 и образования шести молекул 3-ФГА пять из них используются для синтеза рибулозо-5-фосфата, а одна молекула 3-ФГА – для образования глюкозы.
Триозофосфатизомераза превращает 3-ФГА в фосфодиоксиацетон. Затем альдолаза образует из 3-ФГА и фосфодиоксиацетона фруктозо-1,6-дифосфат. Он теряет один остаток фосфорной кислоты под влиянием фруктозо-1,6-дифосфатазы и превращается во фруктозо-6-фосфат. Транскетолаза переносит гликолевый альдегид от фруктозо-6-фосфата на 3-ФГА с образованием эритрозо-4-фосфата и ксилулозо-5-фосфата. Альдолаза присоединяет фосфодиоксиацетон к эритрозо-4-фосфату с образованием седогептулозо-1,7-дифосфата. Он дефосфорилируется фосфатазой и под влиянием транскетолазы соединяется с 3-ФГА. Продуктами этой реакции являются ксилулозо-5-фосфат и рибозо-5-фосфат. Две молекулы ксилулозо-5-фосфата при участии рибулозофосфатэпимеразы и одна молекула рибозо-5-фосфата под действием рибозофосфатизомеразы превращаются в три молекулы рибулозо-5-фосфата.
Если цикл проходит три раза, то образуется 6 молекул ФГА. Пять из шести молекул триозофосфата образуют три молекулы РУбФ, а шестая молекула ФГА является продуктом ассимиляции СО2. Она может превратиться в хлоропласте в первичный крахмал или снова включиться в цикл, или, выйдя в цитозоль, использоваться для образования сахарозы.
Шестая молекула 3-ФГА используется для синтеза фруктозо-1,6-дифосфата при повторении цикла. Из двух молекул фруктозо-1,6-дифосфата образуются фруктозо-6-фосфат и глюкозо-1-фосфат. Последний, взаимодействуя с уридинтрифосфатом, дает уридинфосфоглюкозу. Она и фруктозо-6-фосфат образуют сахарозофосфат, который после дефосфорилирования превращается в сахарозу. Следовательно, для образования одной молекулы сахарозы необходимо прохождение четырех циклов Кальвина. Крахмал синтезируется из уридинфосфоглюкозы под влиянием амилосинтетазы.
Растения: пшеница, рожь, картофель, одуванчик, мать-и-мачеха, большинство сельскохозяйственных культур.

С4, САМ-пути фотосиснтеза, фотодыхание.
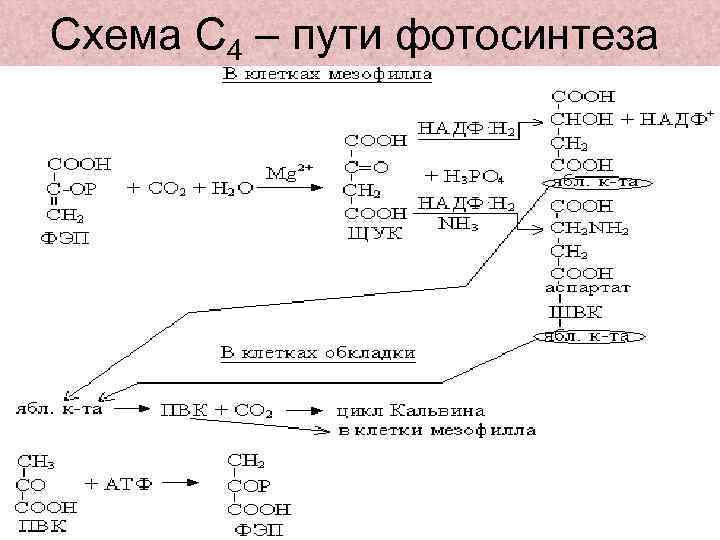
С4-путь фотосинтеза или цикл Хетча-Слэка
Австралийскими учеными М. Хетчем и К. Слэком был описан С4-путь фотосинтеза, характерный для тропических и субтропических растений однодольных и двудольных 16 семейств (сахарный тростник, кукуруза и др.). Большинство самых злостных сорняков – С4 растения, а большинство сельскохозяйственных культур относятся к С3-растениям. Листья этих растений содержат хлоропласты двух типов: обычные в клетках мезофилла и крупные хлоропласты, не имеющие гран и фотосистемы II, в клетках обкладки, окружающих проводящие пучки.
В цитоплазме клеток мезофилла фосфоэнолпируваткарбоксилаза присоединяет СО2 к фосфоэнолпировиноградной кислоте, образуя щавелевоуксусную кислоту. Она транспортируется в хлоропласты, где восстанавливается до яблочной кислоты при участии НАДФН (фермент НАДФ+-зависимая малатдегидрогеназа). В присутствии ионов аммония щавелевоуксусная кислота превращается в аспарагиновую кислоту (фермент - аспартатаминотрансфераза). Яблочная и (или) аспарагиновая кислоты переходят в хлоропласты клеток обкладки, декарбоксилируются до пировиноградной кислоты и СО2. СО2 включается в цикл Кальвина, а пировиноградная кислота переносится в клетки мезофилла, где превращается в фосфоэнолпировиноградную кислоту.
В зависимости от того, какая кислота – малат или аспартат – транспортируется в клетки обкладки, растения делят на два типа: малатный и аспартатный. В клетках обкладки эти С4-кислоты декарбоксилируются, что происходит у разных растений происходит с участием различных ферментов: НАДФ+-зависимой малатдегидрогеназы декарбоксилирующей (НАДФ+-МДГ), НАД+-зависимой малатдегидрогеназы декарбоксилирующей (малик-энзим, НАД+-МДГ) и ФЭП-карбоксикиназы (ФЕП-КК). Поэтому растения делят еще на три подтипа: НАДФ+-МДГ-растения, НАД+-МДГ-растения ФЕП-КК-растения.
Такой механизм позволяет растениям фотосинтезировать при закрытых из-за высокой температуры устьицах. Кроме того, продукты цикла Кальвина образуются в хлоропластах клеток обкладки, окружающих проводящие пучки. Это способствует быстрому оттоку фотоассимилятов и тем самым повышает интенсивность фотосинтеза.
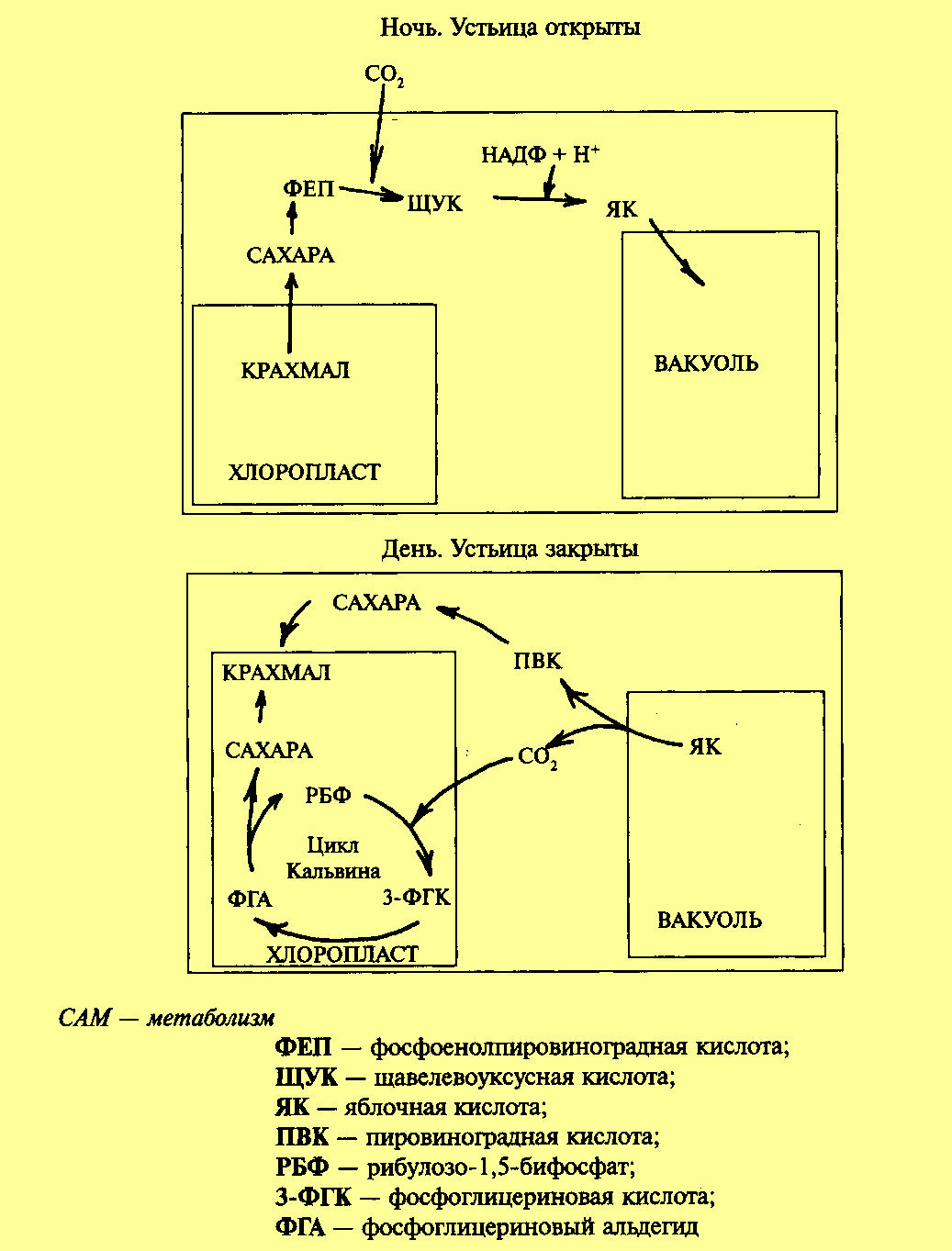
Фотосинтез по типу толстянковых (суккулентов)-САМ путь.
В сухих местах существуют растения-суккуленты, у которых устьица открыты ночью и закрыты днем для уменьшения транспирации. В настоящее время этот тип фотосинтеза обнаружен у представителей 25 семейств.
У суккулентов (кактусов и растений сем. толстянковых (Crassulaceae) процессы фотосинтеза разделены не в пространстве, как у других С4-растений, а во времени. Этот тип фотосинтеза получил название CAM (crassulation acid metabolism)-путь. Устьица днем обычно закрыты, что предотвращает потерю воды в ходе транспирации, и открыты ночью. В темноте СО2 поступает в листья, где фосфоэнолпируваткарбоксилаза присоединяет его к фосфоэнолпировиноградной кислоте, образуя щавелевоуксусную кислоту. Она восстанавливается НАДФН-зависимой малатдегидрогеназой до яблочной кислоты, которая накапливается в вакуолях. Днем яблочная кислота переходит из вакуоли в цитоплазму, где декарбоксилируется с образованием СО2 и пировиноградной кислоты. СО2 диффундирует в хлоропласты и включается в цикл Кальвина.
Итак, темновая фаза фотосинтеза разделена во времени: СО2 поглощение ночью, а восстанавливается днем, из ЩУК образуется малат, карбоксилирование в тканях происходит дважды: ночью карбоксилируется ФЕП, днем РуБФ.
В САМ-растения делят на два типа: НАДФ-МДГ-растения, ФЕП-КК-растения.
Как С4, САМ-тип является дополнительным, поставляющим СО2 в С3-цикл у растений, приспособившихся к жизни в условиях повышенных температур или недостатка влаги. У некоторых растений этот цикл функционирует всегда, у других – только в неблагоприятных условиях.
Фотодыхание.
Фотодыхание – это активируемый светом процесс выделения СО2 и поглощения О2.(НИ К ФОТОСИНТЕЗУ,НИ К ДЫХАНИЮ НЕ ОТНОСИТСЯ). Так как первичным продуктом фотодыхания является гликолевая кислота, оно еще называется гликолатным путем. Фотодыхание усиливается при низком содержании СО2 и высокой концентрации О2 в воздухе. В этих условиях рибулозобисфаткарбоксилаза хлоропластов катализирует не карбоксилирование рибулозо-1,5-дифосфата, а его расщепление на 3-фосфоглицериновую и 2-фосфогликолевую кислоты. Последняя дефосфорилируется с образованием гликолевой кислоты.
Гликолевая кислота из хлоропласта переходит в пероксисому, где окисляется гликолатоксидазой до глиоксиловой кислоты. Образующаяся при этом перекись водорода разлагается каталазой, присутствующей в пероксисоме. Глиоксиловая кислота аминируется, превращаясь в глицин. Глицин транспортируется в митохондрию, где из двух молекул глицина синтезируется серин и освобождается СО2.
Серин может поступать в пероксисому и под действием аминотрансферазы передает аминогруппу на пировиноградную кислоту с образованием аланина, а сам превращается в гидроксипировиноградную кислоту. Последняя при участии НАДФН восстанавливается в глицериновую кислоту. Она переходит в хлоропласты, где включается в цикл Кальвина и образуется 3 ФГА.
Дыхание растений
Живая клетка представляет собой открытую энергетическую систему, она живёт и сохраняет свою индивидуальность за счет постоянного притока энергии. Как только этот приток прекращается, наступает дезорганизация и смерть организма. Энергия солнечного света, запасенная при фотосинтезе в органическом веществе, вновь высвобождается и используется на самые различные процессы жизнедеятельности.
В природе существуют два основных процесса, в ходе которых энергия солнечного света, запасенная в органическом веществе, высвобождается,— это дыхание и брожение. Дыхание — это аэробный окислительный распад органических соединений на простые неорганические, сопровождаемый выделением энергии. Брожение — анаэробный процесс распада органических соединений на более простые, сопровождаемый выделением энергии. В случае дыхания акцептором электрона служит кислород, в случае брожения — органические соединения.
Суммарное уравнение процесса дыхания:
С6Н1206 + 602 -> 6С02 + 6Н20 + 2824 кДж.
Пути дыхательного обмена
Существуют две основные системы и два основных пути превращения дыхательного субстрата, или окисления углеводов:
1) гликолиз + цикл Кребса (гликолитический); Данный путь дыхательного обмена является наиболее распространенным и, в свою очередь, состоит из двух фаз. Первая фаза — анаэробная (гликолиз), вторая фаза — аэробная. Эти фазы локализованы в различных компартментах клетки. Анаэробная фаза гликолиз — в цитоплазме, аэробная фаза — в митохондриях. Обычно химизм дыхания начинают рассматривать с глюкозы. Вместе с тем в растительных клетках глюкозы мало, поскольку конечными продуктами фотосинтеза являются сахароза как основная транспортная форма сахара в растении или запасные углеводы (крахмал и др.). Поэтому, чтобы стать субстратом дыхания сахароза и крахмал должны гидролизоваться с образованием глюкозы.
2) пентозофосфатный (апотомический). Относительная роль этих путей дыхания может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от факторов среды. Процесс дыхания растений осуществляется во всех внешних условиях, при которых возможна жизнь. Растительный организм не имеет приспособлений к регуляции температуры, поэтому процесс дыхания осуществляется при температуре от -50 до +50°С. Нет приспособлений у растений и к поддержанию равномерного распределения кислорода по всем тканям. Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в процессе эволюции разнообразных путей дыхательного обмена и к еще большему разнообразию ферментных систем, осуществляющих отдельные этапы дыхания. При этом важно отметить взаимосвязь всех процессов обмена в организме. Изменение пути дыхательного обмена приводит к глубоким изменениям во всем метаболизме растений.
Энергитическая
11 АТФ образуется в результате работы ЦК и дыхательной и 1 АТФ в результате субстратного фосфорилирования. В ходе этой реакции образуется одна молекула ГТФ (реакция перефосфорилирования приводит к образованию АТФ).
1 оборот ЦК в аэробных условиях приводит к образованию 12 АТФ
Интегративная
На уровне ЦК объединяются пути катаболизма белков жиров и углеводов. цикл Кребса является центральным метаболическим путем, объединяющим процессы распада и синтеза важнейших компонентов клетки.
Амфиболическая
Метаболиты ЦК являются ключевыми на их уровне могут идти переключение с одного вида обмена на другой.