

Сущность фотофизического этапа. Фотохимический этап. Циклический и нециклический транспорт электронов.
Фотофизический этап наиболее важный, т.к. осуществляет переход и преобразование энергии одной системы в другую (в живую из неживой).
Фотофизический этап входит в световую фазу фотосинтеза.
Фотофизический этап начинается с поглощения квантов света, электронами атомов входящих в состав пигментов. В первую очередь кванты света будут поглощаться наиболее подвижными электронами в молекуле хлорофилла, т.е. теми, которые слабее удерживаются ядром (делокализованные p-электроны двойных связей, орбитали которых обобщены между двумя ядрами и неспаренными электронами атомов N2 и О2 в порфириновом ядре). Именно с этим связано то, что молекулы хлорофилла – две основные линии поглощения (в красной и сине-фиолетовой). Из возбужденного первого синглетного и триплетного состояния молекула хлорофилла так же может переходить в основное состояние, при этом ее дезактивация (потеря энергии) может проходить:
1) Путем выделения энергии в виде света или тепла
2) Путем переноса энергии на другую молекулу пигмента
3) Путем затрачивания энергии на фотохимические процессы (потеря электрона и присоединение его к акцептору, с образованием АТФ и НАДФН2)
Молекула пигмента дезактивируется и переходит на основной энергетический уровень.
Магний-порфирин одновременно обладает способностью поглощать и сохранять энергию в виде энергии электронного возбуждения и способностью к окислительно-восстановительным изменениям. Возбужденная молекула хлорофилла – мощный восстановительный агент, играющий решающую роль в образовании высоковосстановленных кофакторов в реакциях фотосинтеза. Основная часть молекул хлорофилла (свето-собирающий комплекс) только поглощает свет и переносит энергию возбуждения на особые молекулы хлорофилла, которые непосредственно учувствуют в фотохимическом процессе. Энергия квантов света улавливается от 200 до 400 молекул антенного хлорофилла свето-собирающего комплекса и стекается к одной молекуле – ловушке, входящей в реакционный центр.
В улавливании и передаче энергии на молекулу хлорофилла-ловушки могут участвовать не только молекулы хлорофилла, но и каротиноиды и фикобиллины. Передача энергии между молекулами пигментов идет главным образом резонансным путем без разделения зарядов с большой скоростью, передача энергии происходит от пигментов, поглощающих свет с меньшей длиной волны к пигментам, поглощающим свет с большей длинной волны. Потеря энергии приводит к превращению квантов более мелкие с большей длиной волны, поэтому основные формы хлорофилла, к которым стекается энергия, являются более длинноволновые, обратный перенос энергии невозможен.
Фотофизический этап заключается в том, что кванты света поглощаются и переводят молекулы пигмента в возбужденное состояние, затем эта энергия приносится на хлорофилл-ловушку, входящую в реакционный центр, осуществляющий первичные фотохимические реакции – разделение зарядов.
Фотохимические реакции фотосинтеза – это реакции, в которых энергия света преобразуется в энергию химических связей, в первую очередь в энергию фосфорных связей АТФ. Именно АТФ обеспечивает течение всех процессов; одновременно под действием света происходит разложение воды, образуется восстановленный НАДФ и выделяется О2.
Энергия поглощенных квантов света стекается от сотен молекул пигментов свето-собирающего комплекса к одной молекуле хлорофилла-ловушке, отдавая электрон акцептору – окисляется. Электрон поступает в электронно-транспортную цепь; предполагается, что свето-собирающий комплекс состоит из 3-х частей: главного антенного компонента и двух фото-фиксирующих систем.
Перенос по цепи переносчиков включает ряд окислительно-восстновительных реакций, при которых происходит перенос либо атома водорода, либо электронов.
В процессе фотосинтеза принимают участие 2 фотосистемы. Сет поглощается раздельно этими системами, и нормальное осуществление фотосинтеза требует их одновременного участия.
Фотосистема 1 – включает светофокусирующие пигменты и реакционный центр 1. Хлорофилл-ловушка 1 поглощает свет длиной волны 700 нм.
Фотосистема 2 – светофокусирующие пигменты и реакционный центр 2. Хлорофилл-ловушка 2 поглощает свет длиной волны 680 нм.
Различают два типа потока электронов: циклический и нециклический.
При циклическом потоке электроны от молекулы хлорофилла передаются к акцептору от молекулы хлорофилла и возвращаются к ней обратно, при нециклическом потоке происходит фотоокисление воды и передача электрона от воды к НАДФ, выделяемая в ходе окислительно-восстановительных реакций энергия частично используется на синтез АТФ.
Подробнее:
В молекуле хлорофилла два уровня возбуждения. Именно с этим связано и то, что он имеет две основные линии поглощения.
Первый уровень возбуждения связан с переходом на более высокий энергетический уровень электрона в системе сопряженных двойных связей, а второй — с возбуждением неспаренных электронов атомов азота и кислорода в порфириновом ядре. При поглощении света электроны переходят в колебательное движение. Наиболее подвижными в молекуле являются делокализованные электроны, орбитали которых как бы размазаны, обобщены между двумя ядрами. Особенно легко возбуждаются электроны сопряженных двойных связей.
Электрон, кроме того, что он находится на определенной орбитали и вращается вокруг ядра, обладает еще спином (вектором магнитного момента) — характеристикой, которую можно трактовать как направление вращения электрона вокруг своей оси. Спин электрона может принимать два значения. Спины двух электронов, находящихся на одной орбитали, противоположны. Когда в молекуле все электроны расположены попарно, их суммарный спин равен нулю. Это основное синглетное состояние (So). В основном энергетическом состоянии So молекула находится в тепловом равновесии со средой, все электроны попарно занимают орбитали с наименьшей энергией. При поглощении света электроны переходят на следующие орбитали с более высоким энергетическим уровнем. При этом имеются две возможности: если электрон не меняет спина, то это приводит к возникновению первого и второго синглетного состояния (S*1, S*2). Если же один из электронов меняет спин, то такое состояние называют
триплетным (Т*1).
Наиболее высокий энергетический уровень — это второй синглетный уровень S*2. Электрон переходит на него под влиянием сине - фиолетовых лучей, кванты которых крупнее, содержат больше энергии. В первое возбужденное S*1 состояние электроны могут переходить, поглощая более мелкие кванты (красного света).
Время жизни на S2 уровне составляет 10~12 с. Это время настолько мало, что на его протяжении энергия электронного возбуждения не может быть использована. Через этот короткий промежуток времени электрон возвращается в первое синглетное состояние S*1 (без изменения направления спина). Переход из второго синглетного состояния S*2 в первое S*1 сопровождается некоторой потерей энергии (100 кДж) в виде теплоты. Время жизни в первом синглетном состоянии немного больше (10~9 или 10~8 с). Наибольшим временем жизни (10~~4-—10~2 с) обладает триплетное состояние Т*1. Переход на триплетный уровень происходит с изменением спина электрона.
Из возбужденного, первого синглетного и триплетного состояния молекула хлорофилла также может переходить в основное. При этом ее дезактивация (потеря энергии) может проходить: 1) путем выделения энергии в виде света (флуоресценция и фосфоресценция – более длительное свечение) или в виде тепла; 2) путем переноса энергии на другую молекулу пигмента; 3) путем затрачивания энергии на фотохимические процессы (потеря электрона и присоединение его к акцептору, образование АТФ и НАДФ-Н2). В любом из указанных случаев молекула пигмента дезактивируется и переходит на основной энергетический уровень. Энергия, испускаемая в виде флуоресценции или в виде теплоты, не может быть использована.
В настоящее время показано, что хлорофилл имеет две функции — поглощение и передачу энергии. При этом основная часть молекул хлорофилла (светособирающий комплекс — ССК) только поглощает свет и переносит энергию возбуждения на особые молекулы хлорофилла, которые непосредственно участвуют в фотохимическом процессе. Такое устройство позволяет значительно полнее использовать энергию света. Подсчитано, что каждая молекула хлорофилла на прямом солнечном свету поглощает квант света не чаще чем через 0,1 с. Между тем скорость последующих реакций фотосинтеза значительно больше. Из сказанного следует, что при непосредственной связи каждой молекулы хлорофилла с последующей реакцией процесс фотосинтеза шел бы прерывисто. Необходимо учитывать также, что использование энергии света в химических реакциях требует большого количества ферментов. Если бы каждая молекула хлорофилла отдавала энергию света непосредственно на фотохимические процессы, то в листе не хватило бы места для размещения всех необходимых для этого ферментных систем. В процессе эволюции в растениях выработался механизм, позволяющий наиболее полно использовать кванты света, падающие на лист подобно каплям дождя. Механизм этот заключается в том, что энергия квантов света улавливается 200—400 молекулами антенного хлорофилла ССК и как бы стекается к одной, особой его молекуле, являющейся ловушкой и входящей в реакционный центр. В улавливании и передаче энергии на молекулу хлорофилла-ловушки могут участвовать не только молекулы хлорофилла, но и каротиноиды и фикобилины.
Передача энергии между молекулами пигментов идет главным образом резонансным путем, без разделения зарядов с большой скоростью. Так, время переноса энергии от одной молекулы хлорофилла к другой составляет 1 х 10 -12—2 х 10 -12 с, а от молекулы каротиноидов к хлорофиллу 4 х 10 -10 с. Таким образом, время переноса энергии значительно меньше времени жизни возбужденной молекулы (10- 8 с). Такой перенос может осуществляться только при близком расстоянии между молекулами пигментов. Расчеты показали, что в одном хлоропласте до 1 млрд. молекул хлорофилла.
Расстояние между молекулами хлорофилла в мембранах составляет всего 1 нм. Перенос энергии происходит только от пигментов, поглощающих свет с меньшей длиной волны, к пигментам, поглощающих свет с большей длиной волны. Дело в том, что хотя передача энергии от одной молекулы пигмента к другой идет с большой эффективностью (от хлорофилла b к хлорофиллу а — 90%, от каротиноидов к хлорофиллу — 40%), однако все же это связано с некоторой ее потерей. Вместе с тем кванты света с меньшей длиной волны обладают большей энергией. Потеря энергии приводит к превращению квантов в более мелкие (с большей длиной волны). Именно поэтому основные формы хлорофилла, к которым стекается энергия, являются более длинноволновыми (хлорофиллы П680 и П700). Обратный перенос энергии невозможен.
Таким образом, в первичных процессах фотосинтеза, связанных с поглощением молекулой хлорофилла кванта света, важную роль играют процессы передачи энергии. Фотофизический этап фотосинтеза и заключается в том, что кванты света поглощаются и переводят молекулы пигментов в возбужденное состояние. Затем эта энергия переносится на хлорофилл-ловушку, входящую в реакционный центр, осуществляющий первичные фотохимические реакции: разделение зарядов.
Дальнейшее превращение энергии света в химическую энергию проходит ряд этапов, начиная с окислительно-восстановительных превращений хлорофилла и включая как фотохимические (световые), так и энзиматические (темновые) реакции.
Циклический транспорт электронов
В первой пигментной системе – пигментом-ловушкой является Р700, а остальные – сборщики (антенный комплекс). При поглощении кванта света один из электронов Р700 переходит на более высокий энергетический уровень и присоединяется к Хл а695 (А0) и филлохинону (А1), а затем белкам, содержащим железо и серу (FeS). Окислительно-восстановительный потенциал Р700 в основном состоянии + 0,43 В; таким образом, тенденция к потере электронов у него выявлена слабо. Когда в результате поглощения кванта света молекула Р700 переходит в возбужденное состояние, ее окислительно-восстановительный потенциал падает до –0,6 В и поэтому она легко отдает электрон (рис. 2.14).
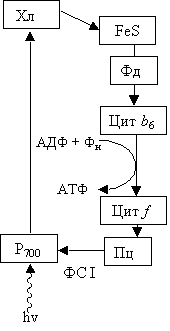
Рис. 2.14. Схема циклического фотофосфорилирования
От белка, содержащего железо и серу, электрон передается переносчику, который называется ферредоксином. От ферредоксина электрон вновь возвращается к Р700 через ряд промежуточных переносчиков, среди которых цитохромы b6 и f, а также пластоцианин. В то время, когда электрон транспортируется по этому циклическому пути, его энергия используется для присоединения Фн к АДФ с образованием АТФ.
Нециклический транспорт электронов
Когда устьица листа открыты, а хлоропласты поглощают свет разных длин волн, одновременно с ФС I включается ФС II. Совместно они составляют систему нециклического транспорта электронов.
Как ранее отмечалось, под действием света Р700 переходит в возбужденное состояние (Р700*) и окисляется (Р700+), электроны переносятся на акцепторы А0 (хлорофилл а) и А1 (филлохинон). Далее на мембрансвязанные железосерные белки (FX, FA, FB), через которые электроны поступают на ферредоксин. Но в этом случае от последнего через флавопротеин – ферредоксин-НАДФ-редуктазу – электроны идут на восстановление НАДФ+ до НАДФН (см. рис. 2.12).

В этом случае от ферредоксина электрон через разные флавопротеиды переходит на НАДФ+. Когда два электрона присоединяются к НАДФ+ из водной среды, которая окружает тилакоидные мембраны, к нему присоединяются протоны воды и образуется НАДФН, который используется в последующих реакциях для восстановления СО2 до уровня углевода.
Таким образом, Р700 окислен, в нем образовалась электронная «дырка». Он не может долго оставаться в этом состоянии. Откуда он получает электрон?
При поглощении кванта света Р680 тоже переходит в возбужденное состояние и отдает электрон первичному акцептору феофитину. При этом хлорофилл Р680 переходит в окисленную форму, а феофитин восстанавливается (Фео –). Затем электрон передается на два хинона QA и QB (менохиноны). После этого электроны поступают на липидрастворимый пул пластохинонов (Q), который функционирует как «двухэлектронные ворота». Один из электронов поступает в цикл цитохромов b6 (цит b/f), а второй направляется по линейной цепи в ФС I. Потом электроны переносятся на водорастворимый белок пластоционин (Пц) и в ловушку Р700 (см. рис. 2.12).
Но теперь «дырка» появилась на Р680. Эта «дырка» заполняется присоединением электрона от воды. Электрон от молекулы воды образуется под воздействием света (фотоокисление или фотолиз). В результате молекула воды разрушается, образуя протоны и кислород:
![]()
Ингибитором этой реакции является диурон (дихлорфенилдиметилмочевина). Растения, обработанные диуроном, гибнут, так как при этом блокируется транспорт электронов от воды, не образуется АТФ и не восстанавливается НАДФ+, а следовательно, невозможно восстановление СО2.
Образованные при фотоокислении воды протоны остаются в водной среде тилакоидов хлоропластов, которая включает не только молекулы воды, но также и Н+- и ОН –-ионы. Накопление протонов на внутренней стороне мембраны тилакоидов играет определенную роль в образовании АТФ, о чем разговор пойдет дальше. Этот путь транспорта электронов получил название нециклического.
Нециклический транспорт электронов также связан с синтезом АТФ. Процесс фосфорилирования АДФ с образованием АТФ, связанный с нециклическим транспортом электронов, получил название нециклического фотосинтетического фосфорилирования (см. рис. 2.12).
![]()
Таким образом, процессы световой фазы фотосинтеза в общем виде можно записать:
Во время световой фазы происходит перенос электронов от воды (Е = +0,81 В) на НАДФ+ (Е = – 0,32 В). Перенос электронов от вещества с более положительным потенциалом к веществу с более отрицательным требует затраты энергии. Используется энергия поглощенного пигментом света. Для транспорта каждого электрона требуется два кванта. Этот перенос электронов против градиента окислительно-восстановительного потенциала и является фотохимической работой.
Следовательно, сущность световой реакции фотосинтеза – вызванный светом последовательный транспорт электронов от одной молекулы к другой, в результате чего образуются АТФ и НАДФН.
Так как молекула, которая теряет электрон, окисляется, а которая получает – восстанавливается, то это окислительно-восстановительный процесс. Для транспорта электронов от одного переносчика ко второму, как и для передачи энергии между молекулами пигментов, необходима тщательная упаковка молекул и строгая упорядоченность в их размещении, поэтому световая стадия фотосинтеза связана с тилакоидными мембранами хлоропластов. Кроме этого, с помощью мембран происходит разобщение процессов образования кислорода и мощного восстановителя, в противном случае, они могли бы взаимодействовать.
Основные различия этих двух видов фосфорилирования даны в следующей таблице (табл. 2.4).
Следовательно, в циклическом транспорте электронов участвует только ФС I, в нециклическом – две. ФС I расположена в тилакоидах стромы, а ФС II – только в тилакоидах гран.
Таблица 2.4. Сравнение циклического и нециклического фотофосфорилирования
Процесс |
Нециклическое |
Циклическое |
Путь электронов |
Нециклический |
Циклический |
Первый донор (источник) электронов |
Вода |
ФС I (Р700) |
Последний акцептор (место назначения) электронов |
НАДФ+ |
ФС I (Р700) |
Продукты |
Основные: АТФ, НАДФН Побочные: О2 |
Основные: АТФ |
Участвующие ФС |
I и II |
I |
Электроны, которые поступают от ферредоксина, могут также переноситься и восстанавливать кислород с образованием Н2О2 и в конечном счете Н2О. В этом случае функционируют ФС I и ФС II, и поглощаются с использованием восстановленного ферредоксина в особой реакции равные количества кислорода; такой поток электронов получил название псевдоциклического. Хотя при этом потоке электронов отсутствует суммарное изменение состояния кислорода, этот процесс нельзя назвать циклическим, поскольку не происходит циклического переноса электронов по указанному пути.
