

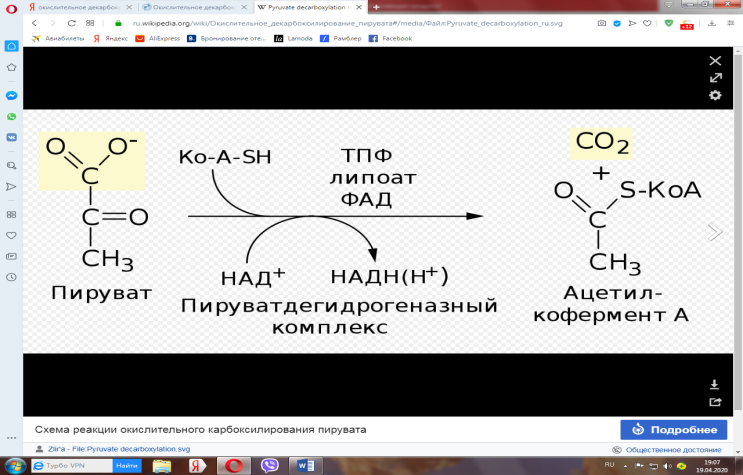
Окислительное декарбоксилирование пировиноградной кислоты
— биохимический процесс, заключающийся в отщеплении одной молекулы углекислого газа (СО2) от молекулы пирувата и присоединении к декарбоксилированному пирувату кофермента А (КоА).
Окисление ПВК до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс». Процесс окислительного декарбоксилирования ПВК происходит в матриксе митохондрий. В нем принимают участие (в составе пируватдегидрогеназного комплекса) 3 фермента:
Пируватдегидрогеназа
Дигидролипоилдегидрогеназа
Дигидролипоилацетилтрансфераза
И 5 коферментов:
тиаминпирофосфат (ТПФ (витамин В1))
амид липоевой кислоты
коэнзим А (СоА (витамин В3))
ФАД (флавинадениндинуклеотид (витамин В2))
НАД+(никотинамидадениндинуклеотид (витамин РР))
Также в состав комплекса ПДК входят два регуляторных белка:
П
 ротеинкиназа
ротеинкиназаФосфопротеинфосфатаза
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
Образовавшийся в процессе окислительного декарбоксилирования ацетил-СоА окисляется далее в цикле трикарбоновых кислот с образованием СО2 и АТФ.
Окислительное декарбоксилирование пирувата — необратимый процесс. Образующийся в ходе этого процесса НАДН впоследствии отдаёт гидридный ион (Н-) в дыхательную цепь, в которой при аэробном дыхании конечным акцептором электронов является кислород, а при анаэробном — другие окисленные соединения (например, сульфат, нитрат). Перенос электронов с НАДН на кислород даёт 2,5 молекулы АТФ на пару электронов.
Стадии:
С
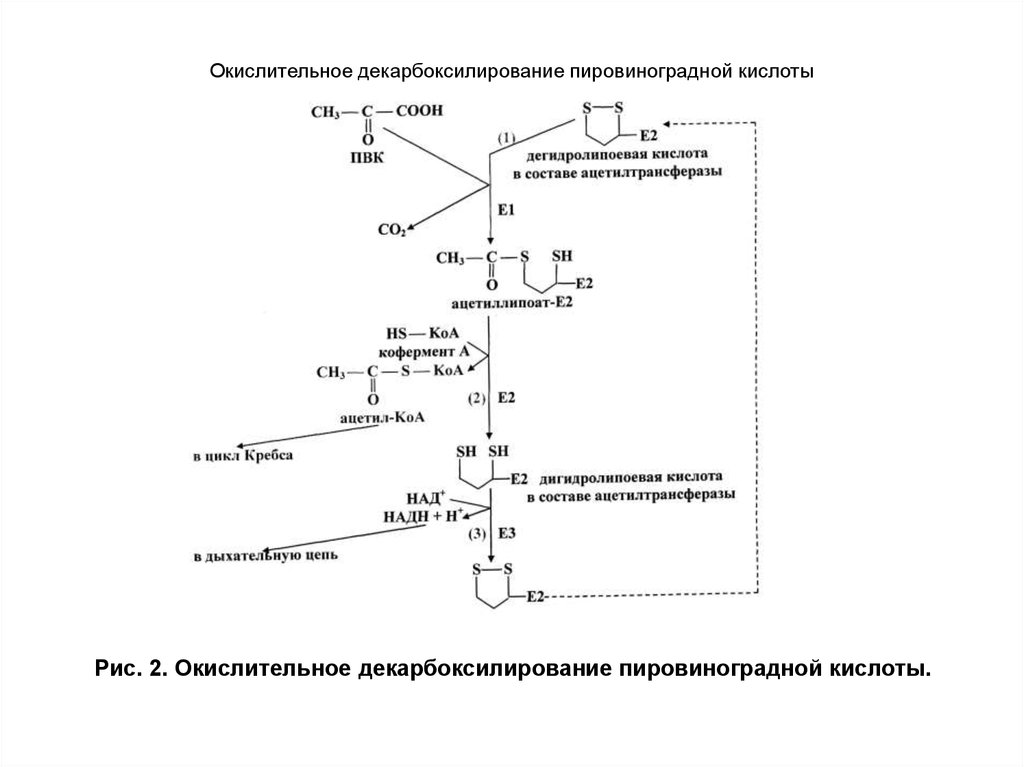 тадия1:
идентична пируватдекарбоксилазной
реакции. Первый атом углерода (С-1)
пирувата уходит в виде СО2, а С-2, в
пирувате находящийся в альдегидной
форме, прикрепляется к ТПФ в виде
гидроксиэтильной группы (—СНОН—СН3).
Первая
стадия является наиболее медленной и
поэтому ограничивает скорость всего
процесса. Кроме того, на этом этапе
комплекс ПДК проявляет свою субстратную
специфичность.
Эта реакция
осуществляется пируватдегидрогеназой
(Е1).
тадия1:
идентична пируватдекарбоксилазной
реакции. Первый атом углерода (С-1)
пирувата уходит в виде СО2, а С-2, в
пирувате находящийся в альдегидной
форме, прикрепляется к ТПФ в виде
гидроксиэтильной группы (—СНОН—СН3).
Первая
стадия является наиболее медленной и
поэтому ограничивает скорость всего
процесса. Кроме того, на этом этапе
комплекс ПДК проявляет свою субстратную
специфичность.
Эта реакция
осуществляется пируватдегидрогеназой
(Е1).Стадия 2: гидроксиэтильная группа окисляется до карбоновой кислоты (ацетата). Два электрона, освобождаемых при этой реакции, идут на восстановление связи —S—S— липоильной группы Е2 до двух тиольных (—SH) групп.
Стадия 3: ацетильный остаток, образующийся в ходе окислительно-восстановительной реакции на стадии 2, сначала связывается тиоэфирной связью с липоильной —SH-группой, а затем переносится на КоА с образованием ацетил-КоА. Таким образом, энергия окисления идёт на образование высокоэнергетического тиоэфира ацетата. Стадии 2 и 3 катализируются дигидролипоилтрансацетилазой (Е2).
Стадия 4 и стадия 5: катализируются дигидролипоилдегидрогеназой (Е3). В ходе этих двух последних реакций восстановленный липоиллизин снова возвращается в окисленную форму, который в дальнейшем может участвовать в следующем цикле окислительного декарбоксилирования пирувата. Электроны, изначально принадлежавшие гидроксиэтильной группе, при этом переносятся с липоиллизина сначала на ФАД с образованием ФАДH2, а потом на НАД+ с образованием НАДН + H+.
Центральную роль в реакции, осуществляемой комплексом ПДК, играют липоиллизиновые «руки» Е2, способные «раскачиваться» и забирать два электрона от Е1, а также ацетильную группу, образовавшуюся из пирувата, и доставлять электроны к Е3. Все эти ферменты и коферменты собраны в комплекс, благодаря чему промежуточные соединения могут вступать в необходимые реакции быстро и не диффундируя с поверхности ферментного комплекса. За счёт этого промежуточные соединения не покидают комплекса, и поддерживается очень высокая локальная концентрация субстрата Е2. Это также предотвращает перехватывание активированной ацетильной группы другими ферментами, использующими её в качестве субстрата.
Биологическая роль определяется тем, какова дальнейшая судьба продуктов: образовавшаяся молекула НАДН2 окисляется в длинной дыхательной цепи с образованием 3-х молекул АТФ. Так же является важным этапом катаболизма, позволяющим включаться в цикл Кребса тем веществам, при распаде которых образуется ПВК.
Цикл Кребса - цепочка химических реакций, происходящих в митохондриях. Является общим конечным путем окисления ацетильных групп (в виде ацетил-КоА).
Когда ацетил-КоА попадает в митохондриальный матрикс, он связывается с молекулой оксалацетата и превращается в лимонную кислоту (цитрат). Цитрат, в свою очередь, под действием фермента аконитазы превращается в цис-аконитат, оставляя молекулу воды. В свою очередь цис-аконитат превращается в изоцитрат под действием фермента изоцитратдегидрогеназы. Изоцитрат превращается в альфа-кетоглутарат под действием изоцитратдегидрогеназы. Альфа-кетоглутарат превращается в сукцинил-КоА альфа-кетоглутаратдегидрогеназой и добавлением ацетил-КоА. Он подвергается сукцинату под действием сукцинат-тиокиназы, которая превращает его в фумарат. Фумарат превращается в L-малат через фумаразу. L-малат под действием фермента малатдегидрогеназы восстанавливает оксалацетат, который может снова вступать в реакцию с молекулой ацетил-КоА и повторять цикл.
Результатом этого цикла является образование CO2 и водорода, а также воды. Ионы на выходе из процесса участвуют в ресинтезе АТФ.
Окисление ацетильного остатка происходит в несколько стадий, образующих циклический процесс из 8 основных этапов:
К
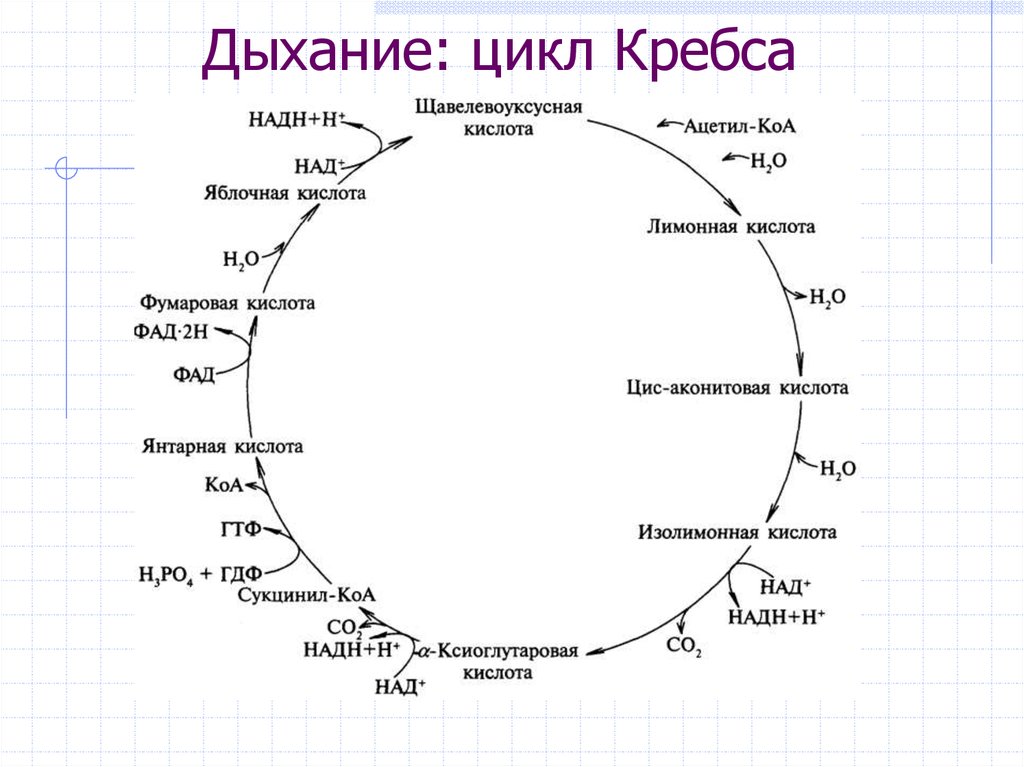 онденсация
ацетил-КоА и оксалоацетата с образованием
цитрата: происходит реакция отщепление
карбоксильной группы аминокислот, в
процессе которой образуется ацетил-КоА,
он выполняет функцию транспортировки
углерода в различных обменных
процессах.
При соединении с молекулой
щавелевой кислоты получается цитрат,
который фигурирует в буферных обменах.
На
данном этапе кофермент А полностью
высвобождается, и получается молекула
воды.
Реакция необратима.
онденсация
ацетил-КоА и оксалоацетата с образованием
цитрата: происходит реакция отщепление
карбоксильной группы аминокислот, в
процессе которой образуется ацетил-КоА,
он выполняет функцию транспортировки
углерода в различных обменных
процессах.
При соединении с молекулой
щавелевой кислоты получается цитрат,
который фигурирует в буферных обменах.
На
данном этапе кофермент А полностью
высвобождается, и получается молекула
воды.
Реакция необратима.Превращение цитрата в изоцитрат: дегидрирование (отщепление молекул воды) от цитрата, в результате которого получается цис-аконитат. Он присоединяя молекулу воды, переходит в изолимонную кислоту (изоцитрат).
Превращение изоцитрата в а-кетоглутарат: изолимонная кислота(изоцитрат) дегидрируется в присутствии НАД- зависимой изо-цитратдегидрогеназы.На выходе получаем альфа-кетоглутарат, который участвует в регуляции всасывания аминокислот, нормализует метаболизм и положительно влияет на антистрессорные процессы. Образуется NADH (аллостерический фермент).
Окисление α-кетоглутарата до сукцинил-КоА: окислительное декарбоксилирование а-кетоглутарата с образованием сукцинил-КоА - тиоэфира, содержащего высокоэнергетическую фосфатную связь.
Превращение сукцинил-КоА в сукцинат: пятая реакция катализируется ферментом сукцинил-КоА-синтетазой. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Происходит образование высокоэргической фосфатной связи ГТФ за счет тиоэфирной связи сукцинил-КоА.
Дегидрогенирование сукцината, образование фумарата: образовавшийся сукцинат превращается в фумарат под действием фермента сукцинат-дегидрогеназы. Единственная дегидрогеназная реакция цикла Кребса, в ходе которой осуществляется прямой перенос водорода с субстрата на флавопротеин без участия НАД+.
Образование малата из фумарата: под влиянием фермента фумаратгидратазы (фумаразы). Образовавшаяся при этом фумаровая кислота гидратируется, продуктом реакции является L-яблочная кислота (L-малат).
Превращение малата в оксалоацетат: под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат. Происходит полное «сгорание» одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА. А коферменты (НАД+ и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться.
Значение:
Энергетическое:
В цикле Кребса выделяют 3 реакции, идущие с образованием НАДН2, их катализируют ферменты изоцитратдегидрогеназа, кетоглютаратдегидрогеназный комплекс, малатдегтдрогеназа. Образовавшиеся в ЦТК 3 молекулы НАДН2в последующем окисляются в длинной ЦПЭ с образованием 9 молекул АТФ (при окислении каждой НАДН2синтезируется 3 молекулы АТФ).
Образовавшийся в ЦТК ФАДН2 окисляется в короткой ЦПЭ, давая энергию для синтеза 2 молекул АТФ.
В сукцинилтиокиокиназной реакции ЦТК непосредственно образуется 1 макроэрг – ГТФ (1 ГТФ = 1 АТФ).
В целом общая энергетическая эффективность ЦТК составляет 12молекул АТФ.
Анаболическое: некоторые метаболиты цикла Кребса не окисляются в нём, а используются для синтеза новых веществ. Например:
α - кетоглютаровая используется на синтез глютаминовой кислоты.
Сукцинил-КоА используется на синтез гема.
Ацетил КоА идёт на синтез жирных кислот, холестерина.
Щавелевоуксусная кислота может участвовать в синтезе аспарагиновой кислоты.
Интегративное: объединяет пути катаболизма углеводов, жиров, белков, во всех аэробных организмах он выступает в роли центрального метаболического пути углерода.