

ГЕНЕТИКА И СЕЛЕКЦИЯ
ВВЕДЕНИЕ В ГЕНЕТИКУ
Генетика – наука о закономерностях наследственности и изменчивости.
Наследственность – присущее всем организмам свойство обеспечивать в ряду поколений преемственность признаков и особенностей развития.
Изменчивость – свойство организмов изменять свои признаки и свойства, обусловливающее разнообразие индивидов, популяций, рас и т.д.
Краткая история генетики
Античность:
Теория прямого наследования признаков |
Теория непрямого наследования признаков |
Гиппократ |
Аристотель |
Репродуктивный материал собирается из всех частей тела и таким образом все органы тела непосредственно влияют на признаки потомства |
Репродуктивный материал производится из питательных веществ, по своей природе предназначенных для построения разных частей тела |
Теория пангенезиса Дарвина (1868 г.) |
Ф. Гальтон (1871 г.) |
Труды Г. Менделя
1865 г. – «Опыты над растительными гибридами»
Законы непрямого наследования
Стали основой генетики
Переоткрытие законов Менделя
1900 г. – К. Корренс, Э. Чермак, Г. Де Фриз – растения
1902 г. – У. Бэтсон, Л. Кюэно – животные
1889 г. – Т. Бовери, ядерная гипотеза наследственности
1903 г. У. Сэттон – параллелизм в поведении менделевских факторов и хромосом
А. Вейсман – невозможность наследования признаков, приобретённых в онтогенезе, автономия зародышевых клеток
Мутационная теория
1901 г. – Г. Де Фриз
1899 г. – С. И. Коржинский, теория гетерогенеза
Наследственные признаки могут скачкообразно изменяться вследствие мутирования
Хромосомная теория наследственности
Т. Х. Морган и его школа: А. Стёртевант, К. Бриджес и Г. Мёллер
Drosophila melanogaster
Линейное расположение генов в хромосомах
Ген – элементарный носитель наследственной информации
1920 г. – Н. И. Вавилов, закон гомологических рядов наследственной изменчивости
Открытие индуцированного мутагенеза
1925 г. – Г. А. Надсон и Г. С. Филиппов – дрожжи
1927 г. – Г. Мёллер – дрозофила
1927 г. – Дж. Стадлер – растения
Исследования по генетике в СССР
1919 г. – Ю. А. Филипченко, первая кафедра генетики, Петроградский университет
1929 г. – первый учебник «Генетика»
1930 г. – А. С. Серебровский – кафедра генетики, Московский университет
Институт экспериментальной биологии, Москва, Н. К. Кольцов
Рождение молекулярной генетики
Джордж Билд, Эдвард Тейтум: один ген – один фермент
1944 г. – О. Эвери, К. Маклеод, М. МакКарти – генетическая роль нуклеиновых кислот
1953 г. – Дж. Уотсон, Ф. Крик – расшифровка структуры молекулы ДНК
1958 г. – М. Мезельсон, Ф. Сталь – полуконсервативный характер удвоения ДНК
1961 г. – Ф. Крик: генетический код состоит из триплетов
1966 г. – Р. Холли, Х. Корана и М. Ниренберг – расшифровка генетического кода
Рождение генной инженерии
1972 г. – в лаборатории Берга была создана первая рекомбинантная ДНК
1970-80-е – методы копирования и расшифровки ДНК in vitro (полимеразная цепная реакция (ПЦР) и секвенирование)
Геномная эра
1977 г. – секвенирован геном бактериофага Ф-Х174
1995 г. – секвенирован геном бактерии Haemophilus influenza
1996 г. – секвенирован геном пекарских дрожжей Saccharomyces cerevisiae
1998 г. – секвенирован геном нематоды Caenorhabditis elegans
2001 г. – первые результаты проекта «Геном человека»
2003 г. – Проект «Геном человека» успешно завершён
Попытки искусственной сборки геномов
Значение генетики для других наук и практики
Генетика и селекция
Различные системы скрещиваний
Гибридологический анализ
Индуцирование мутаций, полиплоидизация
Клеточная и генная инженерия
Гибридизация соматических клеток растений
Генетика и медицина
Генные мутации и хромосомные аберрации вызывают целый ряд аномалий обмена веществ, нарушений конституции и психических заболеваний
Ранняя диагностика
Медико-генетическое консультирование
Генотерапия
Генетика и экология
Генетика популяций
Сохранение генофондов
Экологическая генетика
Изучение мутагенной активности разнообразных физических и химических агентов
Испытание на генетическую активность с помощью тест-систем
Генетика и другие биологические науки
Генетика и биохимия – молекулярная биология
Генетика и дарвинизм – синтетическая теория эволюции
Генетика животных, растений, микроорганизмов – зоология, ботаника, микробиология
Поведенческие мутанты – физиология животных, физиология высшей нервной деятельности
Биохимические мутанты – биохимия
МАТЕРИАЛЬНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ
ДНК – носитель наследственной информации
ДНК – полидезоксирибонуклеотид (полинуклеотид)
Мономеры – дезоксирибонуклеотиды
Нуклеотиды – фосфорные эфиры нуклеозидов (нуклеозидтрифосфаты)
Нуклеотид – азотистое основание + пентоза (дезоксирибоза) + остаток фосфорной кислоты
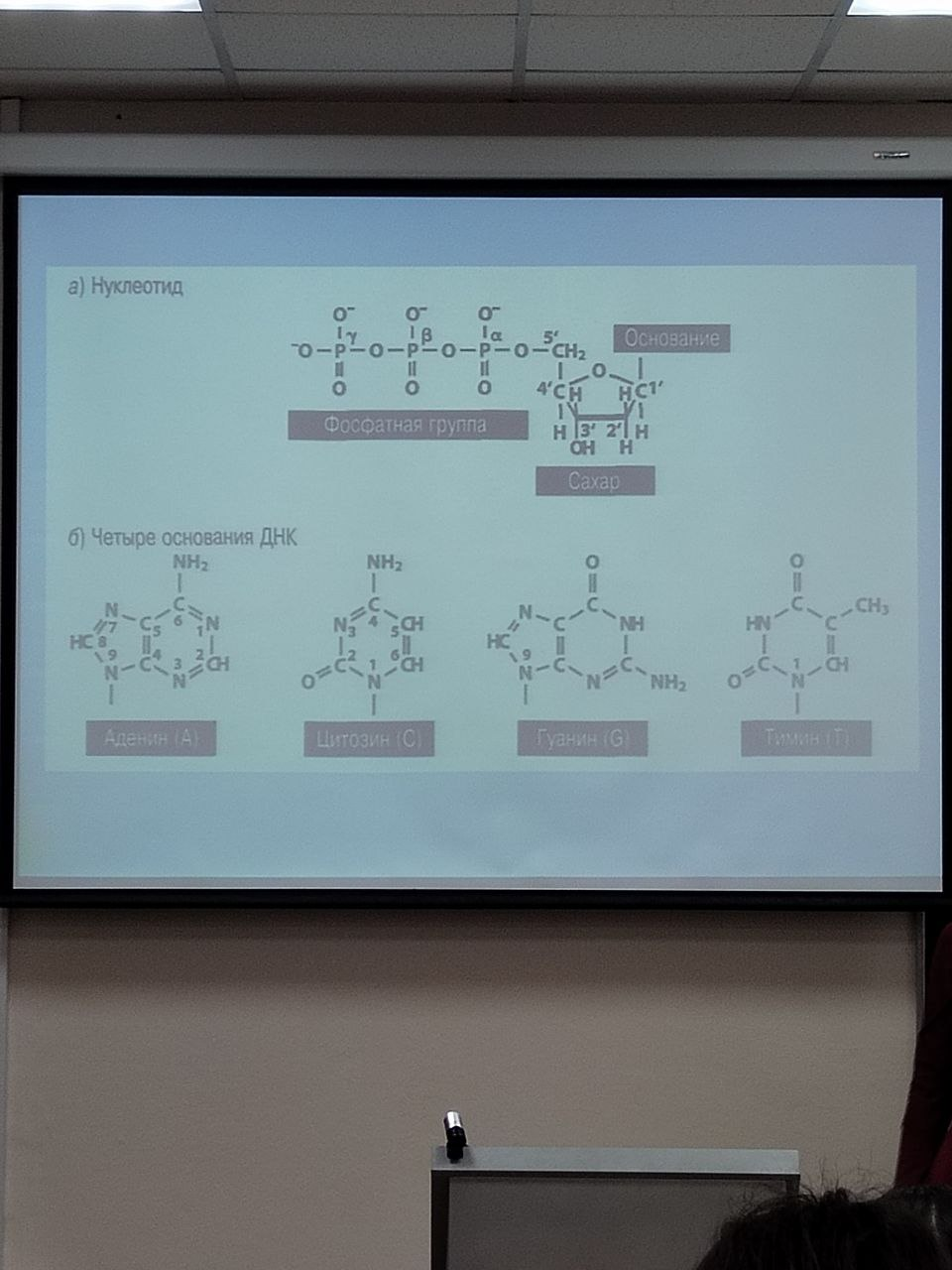
Правило Чаргаффа: A=T, G=C

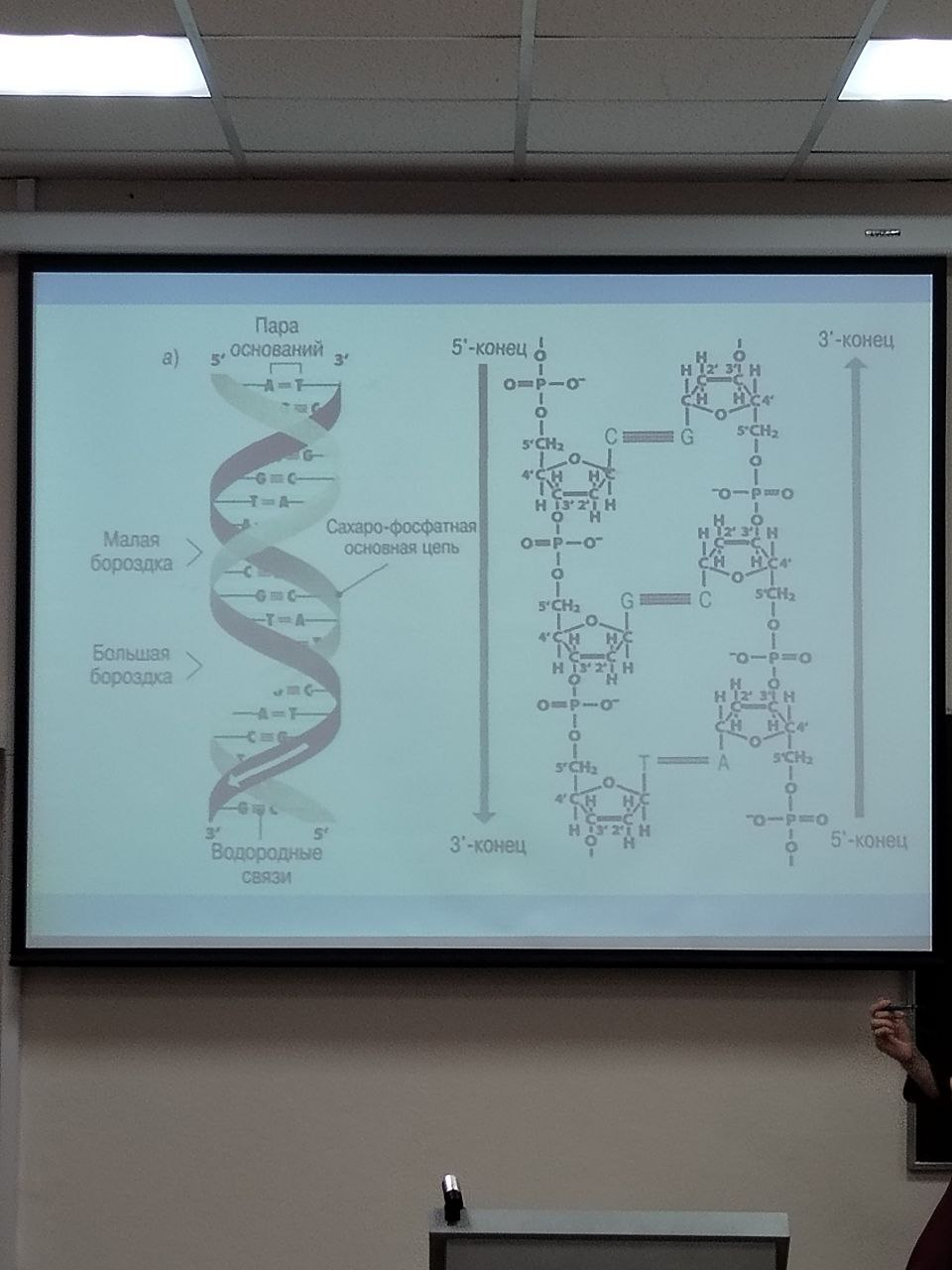
Молекулярная модель ДНК
Число полинуклеотидных цепей равно двум
Цепи образуют правозакрученные спирали по 10 оснований в каждом витке
Цепи закручены одна вокруг другой и вокруг общей оси
Цепи антипараллельны
Фосфатные группировки находятся снаружи спиралей, а основания – внутри и расположены с интервалом 0,34 мкм под прямым углом к оси молекулы
Цепи удерживаются вместе водородными связями между основаниями
Пары, образуемые основаниями А-Т и G-C, в высшей степени специфичны
Полинуклеотидные цепи комплементарны друг другу
Гены отличаются чередованием пар нуклеотидов
Наследственная информация закодирована в виде последовательности нуклеотидов
Мутации – результат изменения чередования нуклеотидов
Воспроизведение генов обеспечивается комплементарностью оснований нуклеотидов – происходит за счёт разъединения комплементарных полинуклеотидных цепей и достройки новых, комплементарных цепей из нуклеотидов клетки.
Конвариантная редупликация
Генетическая роль днк
Трансформация бактерий
1928 г. – Ф. Гриффит, эксперименты со стрептококком
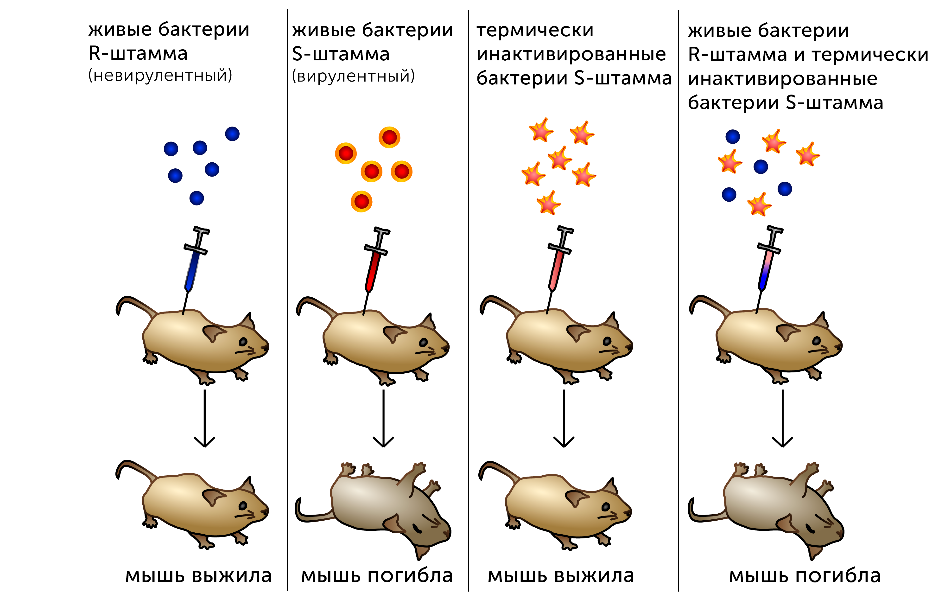

1944 г. – О. Эвери, К. МакЛеод и М. МакКарти идентифицировали трансформирующий агент как ДНК
Дезоксирибонуклеазы (ДНКазы) приводят к потере способности обработанных ими клеток вызывать трансформацию
Размножение бактериофагов
1952 г. – А. Херши и М. Чейз, эксперимент с бактериофагом Т2

Сопоставление плоидности и содержания ДНК в клетке
1949 г. – Х. Рис, А Мирский

Видовая специфичность нуклеотидного состава ДНК
G+C/A+T – коэффициент нуклеотидной (видовой) специфичности
Геносистематика – сравнение состава и структуры нуклеиновых кислот для построения естественной системы организмов
Генетический код
1961 г., Ф. Крик – «Общая природа генетического кода для белков»
Код триплетен (43=64)
Код не перекрывается
Код вырожден, т.е. избыточен, имеет синонимы (часто у таких кодонов совпадают первые две позиции)
Между кодонами нет «запятых»
Считывание кода в пределах гена происходит с фиксированной точки в одном направлении
Код универсален для всех живых организмов

РНК как генетический материал
Вирус табачной мозаики (ВТМ)

Гены и их экспрессия
Ген – функциональная единица наследственности
Каждый ген – последовательность нуклеотидов, несущая инф-ю о конкретном продукте (РНК или белке).
Ген – дискретный фактор, передаваемый в неизменном виде от родителей потомству.
Гипотеза «один ген-один фермент»
Бидл и Тейтум: каждый шаг метаболизма катализируется определенным ферментом, за продукцию которого ответственен один ген, и мутация в этом гене влияет на активность закодированного фермента. Есть исключения!
Один ген – одна полипептидная цепь!
Ген – участок ДНК, который транскрибируется в РНК-копию по одной из нитей ДНК.
Большинство генов несут инф-ю о последовательности АК-остатков в белке, однако некоторые гены кодируют только РНК.
Иногда регуляторные последовательности ДНК иногда тоже называют генами.
Экспрессия генов – совокупность процессов, приводящих к синтезу функционального продукта гена. Объединяет процессы транскрипции, трансляции, регуляцию этих процессов.
Чаще всего регуляция экспрессии генов осуществляется на уровне транскрипции.
Гены расположены на хромосомах
Хромосома содержит линейно расположенные на ней гены;
Каждый ген занимает на хромосоме строго определенное положение – генетический локус;
Аллели гена – это слегка отличающиеся варианты последовательности, находящиеся в данном генетическом локусе.
Мутации генов
Мутация – случайное событие, связанное с изменением структуры гена. Ненаправленное изменение.
Вероятность того, что мутация нарушит или даже аннулирует функцию гена, достаточно высока;
Большинство мутаций, влияющих на функцию генов, рецессивны;
Доминантная аллель – это та, которая обеспечивает наработку достаточного количества активного белка.
Тест на аллелизм
Для того чтобы понять, находятся ли мутации, приводящие к одному фенотипу, в одном гене или в разных, используют комплементирующий тест. Получают гетерозиготное по обеим мутациям поколение организмов.
Влияние мутаций на фенотип
Мутация идет с потерей функции – рецессивна
А) нуль-мутация – белок не синтезируется;
Б) мутация с нарушением функции;
Мутация с приобретением функции – доминантна
Молчащая мутация – слабая мутация
Нуклеотидные замены не приводят к изменениям в АК-составе белка;
Изменение АК-состава не приводит к изменению активности белка.
Множественный алеллизм
Множественные аллели – разные варианты одного и того же гена.
Взаимодействие между аллелями может принимать различные формы.
У конкретного генетического локуса может быть более одного аллеля дикого типа.
Группы крови системы АВ0 у человека
Локус АВ0 кодирует фермент галактозил-трансферазу.
Полиморфизм локуса – три варианта фермента, отличающиеся способностью взаимодействовать с разными кофакторами
Аллели А и В доминантны.
В случае, когда в популяции присутствуют множественные функциональных аллелей одного гена обозначают генетический полиморфизм.
Прокариотические гены
Колинеарность гена и белка – линейное соответствие между последовательностью нуклеотидов в гене и последовательностью АК в белке.
Доказана для бактерий и вирусов;
Ген, состоящий из 3N пар нуклеотидов, должен состоять из N АК.
Эукариотические гены
У эукариот в пределах кодирующего региона могут находиться дополнительные последовательнсоти, прерывающие участок, кодирующий тот или иной белок.
Гены эукариот состоят из экзонов и интронов.
Экзоны – участки ДНК, из которых состоит зрелая мРНК. По определению, ген начинается и заканчивается экзонами, соответсвующими 5-3 кончами РНК
Интроны – внутренние регионы гена, удаляемые из первичного транскрипта
Ген длиннее, чем белок-кодирующая последовательность!
Элементы гена:
Промотор – участок ДНК, контролирующий экспрессию гена, может быть довольно отдален от гена;
Точка начала транскрипции TSS;
Лидерная нетранслируемая область leader UTR
Старт-кодон
Кодирующий регион (экзоны и интроны)
Стоп-кодон
Трейлерная нетраслируемая область trailer UTR
Точка окончания транскрипции TES.
Ген не напрямую транслируется в белок, а посредством матричной РНК.
У бактерий экспрессия генов происходит в две стадии, происходящие в одном и том же клеточном компартменте:
Транскрипция – с одной из цепей ДНК синтезируется копия мРНК
Трансляция – последовательность мРНК, состоящая из триплетов, кодирующих АКЮ формирует соотвествующий белок.
У эукариот в интросодержащих генах синтезуируется более длинный предшественник мРНК – пре-мРНК. Модификация пре-мРНК, происходящие после ее синтеза на ДНК – процессинг РНК. Процессы происходят в ядре.
Модификации пре-мРНК:
Сплайсинг – удаление интронов из пр-мРНК;
Модификация 5штрих и 3 штрих концов --- зрелая мРНК, содержащая протяженную открытую рамку считывания.
Некоторые последовательности ДНК кодируют более одного белка
Перекрывающиеся гены – возможно образование продуктов разной длины (полноразмерная и укороченная форма белка)
Последовательность ДНК принимает участие в синтезе двух негомологичных белков (сдвиг рамки считывания)
Альтернативный сплайсинг.
Длина генов
Переключение с преимущественно безинтронных генов на преимущественно интронные гены происходит у низших эукариот.
Увеличение длины генов происходит преимущественно за счет интронов.
С увеличением длины гена корреляция между размером генома и сложностью организации вида исчезает. С увеличением размера генома в первую очередь увеличиваются размеры интронов, а экзоны остаются прежних размеров – довольно короткие. Т.е. гены эволюционируют путем добавления участков, кодирующих небольшие отдельные домены белков. Размер гена в первую очередь зависит от длины его интронов.
Наличие интронов дает возможность альтернативного сплайсинга и повышает эволюционную пластичность вида.
Матричные процессы
Матричными называются такие процессы, при которых на основе первичной структуры одного биополимера осуществляется синтез другого биополимера, называемой матрицей, синтезируется первичная структура другого биополимера, называемого копией, причем структура матрицы определяет структуру копии.
К матричным процессам относятся:
Биосинтез ДНК, или репликация
Биосинтез РНК, или транскрипция
Биосинтез белка, или трансляция.
Любой матричный процесс можно разбить на 3 фазы:
Начало синтеза, или инициация
Продолжение синтеза, или элонгация
Окончание синтеза, или терминация
Ферментативные процессы
Энергозависимые процессы
Требуют затрат неферментных белковых факторов
Мутации генов, ответственных за матричные процессы, имеют плейотропное действие, как правило, летальные
Ведущим правилом всех матричных процессов является правило комплементарности.
Репликация ДНК
Полуконсервативный механизм репликации ДНК
1957 г – эксперимент Мезельсона и Сталя доказал полуконсервативный механизм репликации.
Механизм репликации
Реплицируется целая молекула ДНК перед каждым клеточным делением. «Репликационная вилка» - особая четко ограниченная область репликации которая перемещается вдоль родительской ДНК.
В составе репликативной вилки выделяют две реплисомы – мультиэнзимные комплексы ферментов, ответственные за репликацию ДНК.
Репликоны – единица репликации, в пределах которой она начинается и заканчивается
У прокариот обычно образуется одна репликативная вилка, которая движется в противоположных направлениях, пока они
У эукариот образуются множество репликативных вилок, которые движутся до тех пор пока не встретятся друг с другом.
Инициация
Ориджин репликации - точка старта:
OriC у прокариот, богатый АТ-парами.
Несколько более сложных ориджинов у эукариот.
Необходимо раскручивание ДНК с образованием однонитевых фрагментов ДНК в стабильном состоянии (SSB белки), а также сброс возникающих супервитков (ферменты топопизомеразы).
Для начала репликации необходимы инициирующие белки (DnaA у прокариот; ORC Cdk Ddk у эукариот) и ферменты хеликазы для плавления ДНК (разделения двух цепей).
Элонгация
Главный фермент – ДНК-зависимая-ДНК полимераза в направлении 5-3 (растущая цепь), при этом по матричной цепи фермент движется в направлении 3-5.
Цепи ДНК антипараллельны – репликационная вилка ассиметрична: одна из дочерних цепей ДНК строится непрерывно (лидирующая, ведущая), а другая непрерывна (отстающая).
Наращивание второй цепи идет отдельными фрагментами – фрагментами Оказаки. Для их соединения нужны ДНК-лигазы. Образование каждого фрагмента Оказаки возможно тогда, когда репликативная вилка сдвинется и откроется соответствующий участок матрицы.
ДНК-полимераза способна только добавлять новые основания к уже имеющемуся 3-ОН концу цепи. Эту преобразованную цепь называют праймером или затравкой. Наличие затравки обеспечивается праймазой, которая синтезирует короткий РНК-праймер, который ы дальнейшем будет удален при репликации.
Эндонуклеазы – удаление РНК-затравок; особый вид полимеразы заполняет образовавшиеся пустоты.
Корректорские функции ДНК полимеразы – обнаруживание неправильно присоединенных нуклеотидов.
В ходе репликации с небольшой частотой ДНК-полимеразы могут допускать ошибки и присоединять неправильные нуклеотиды.
Для координации репликационного комплекса необходимо близкое расположение ферментов синтеза ДНК на обоих цепях, что достигается путем выпетливания одноцепочечных участков на отстающей цепи («тромбонная петля»).
Терминация
У прокариот имеется определенный участок, на котором заканчивается репликация ДНК TerС
У эукариот завершение происходит когда сливаются репликативные вилки.
Концевые участки хромосом представлены особыми участками, называемыми теломерами. После прохождения цикла репликации и укорочения теломер в эукариотах включается особый ферментный комплекс – теломераза, чтобы предотвратить потенциально укорочение смысловой части на концах хромосом. Лимит Хейфлика – ограничение числа клеточных делений, связанное с укорочением теломерных участков.
Транскрипция ДНК
Транскрипцией называется перенос информации с двуцепочечной молекулы ДНК на одноцепочечные молекулы РНК
Матрицей для синтеза РНК служит только одна цепь ДНК – матричная цепь.
ДНК-зависимая РНК-полимерааза. Направление от5 к 3 концу растущей цепи РНК.
Инициация
РНК-полимераза прокариот состоит из 4 субъединиц 2 альфа и 2 бета цепи – минимальный фермент не способен «узнавать» на ДНК специфические стартовые позиции начала транскрипции – промоторы.
Сигма-фактор – дополнительная субъединица для узнавания промоторов.
Нуклеотидные последовательности промоторов – ТАТА-бокс или бокс Прибнова.
Для эукариот необходимы особые белки – факторы транскрипции, привлекающие РНК-полимеразу. TTS и TES сайты.
Элонгация
Производится ферментов РНК-полимеразой без сигма-фактора. У прокариот РНК-полимераза способна синтезировать все типы молекул клеточной РНК.
У эукариот известны три типа РНК полимераз:
1 – синтез рРНК
2 – синтез мРНК
3 – синтез тРНК
Терминация
Может происходить 2 путями:
У прокариот может происходить в специальных участках ДНК – терминаторах, содержащих инвертированные повторы – палиндромы, или шпильки
У прокариот и эукариот есть также Rho-зависимый механизм терминации, требующий наличие Rho-белка.
Только небольшая фракция клеточной РНК в каждый данный момент отражает состав ДНК. Регуляция транскрипции в основном проходит на уровне инициации транскрипции за счет транскрипционных факторов, которые могут как подавлять (репрессоры) так и активировать транскрипцию.
У эукариот существуют также активирующие и репрессирующие регионы, расположенные на большом удалении от точки начала транскрипции (энхансеры и сайленсеры). Модификации гистоновых белков также регулируют транскрипцию эукариот.
Трансляция мРНК
Синтез белка на матрице мРНК. Информация переводится с четырехбуквенного алфавита НК на 20-тибуквенный алфавит АК-последовательностей полипептидных цепей.
В процессе трансляции ключевым фактором является рибосома.
Рибосома – молекула рРНК, ассоциированная с белками. Белки – особые белковые факторы трансляции. Состоит из 2 субъединиц, большой и малой. Коэффициент седиментации у прокариот 70S (16S и 23S); у эукариот 80S (18S и 28S). Обладают крайне высокой видовой специфичностью
Малая субъединица – инициация трансляции. Большая субъединица – элонгация.
тРНК – устанавливает соответствие между кодоном и АК. Структура тРНК «клеверный лист»: акцепторный стебль, антикодоновая петля.
Три стадии транспорта АК:
Стадия активации АК – образование аминоациладенилатов
Стадия аминоацилирования тРНК – присоединение АК-остатков к тРНК
Собственно трансляция, или полимеризация АК-остатков с образованием пептидных связей
Инициация начинается со старт-кодона АУГ, что обеспечивается тРНК. У прокариот последовательность Шайна-Дальгарно обеспечивает взаимодействия мРНК с рРНК.
Элонгация: 3 сайта в составе рибосом. молекула тРНК подходит к А-сайту, который соответствует одному кодону. тРНК перемещается в Р-сайт, в котором происходит образование пептидной связи. Далее пустая тРНК попадает в Е-сайт и вытесняется оттуда новой тРНК.
Терминация происходит, когда рибосома доходит до стоп-кодона. В этот момент в А-сайт попадают белковые факторы терминации трансляции.
Репарация ДНК. Репликация
Системы репарации – ферментные системы, распознающие повреждения ДНК и корректирующие их.
Системы репарации по сложности сравнимы с репликативным аппаратом, важны для выживания клетки.
Когда репаративная система исправляет изменение ДНК, оно не имеет последствий. Если системе это не удается, результатом может быть мутация.
Измеряемая скорость возникновения мутаций отражает баланс между накоплением повреждений ДНК и скоростью их исправлений.
Типы систем репарации:
Прямое исправление повреждений ДНК
Репарация азотистых оснований или целых нуклеотидов
Репарация путем рекомбинации
Путь негомологичного соединения концов
Мутации в генах, отвечающих за систему репарации, часто приводят к развитию онкологических заболеваний.
Типы повреждений ведущих к запуску систем репарации:
Однонуклеотидные замены
Нарушают нуклеотидную последовательность, но не структуру ДНК в целом
Не влияют на процессы транскрипции или репликации, но имеют эффект у будущих поколений клеток
Некомплементарность сохраняется только до следующей репликации.
Искажения структуры ДНК
Могут создавать физические помехи при репликации или транскрипции
Вредоносная химическая модификация будет оставаться в ДНК и вызывать структурные дефекты и/или провоцировать мутации до тех пор, пока не будет удалена.
Виды репарации
Прямая репарация
Повреждения просто удаляется или исправляется на месте. Встречается редко. Пример, фотореактивация пиримидиновых димеров фотолиазой.
Эксцизионная репарация
Включает узнавание поврежденного участка и его вырезание и репаративный синтез ДНК специальной ДНК-полимеразой. Используется для:
Исправления химических модификаций оснований;
Устранения некомплементарности ДНК из-за неспаренных оснований (мисмэтч репарация). Требует способности отличать старую и новую цепи ДНК – исправления вносятся в новую цепь. Пример, метилирование старых цепей ДНК – метилазы.
Два подтипа:
- репарация с эксцизией оснований BER – система напрямую вырезает из ДНК поврежденное азотистое основание и заменяет его; пример, урацилгликозилаза
- репарация с эксцизией целиком NER – система вырезает из ДНК последовательность =, содержащую поврежденное основание; затем на замену на замену вырезанному материалу синтезируется новый отрезок ДНК.