

1.Введение в фармакологию ЦНС
.pdf20
Пресинаптическое торможение:
Пресинаптическое торможение – осуществляется благодаря тому, что с возбуждающим аксоном заранее
проконтактирует вставочный тормозной нейрон → аксон не успеет высвободить нейромедиатор и провести возбуждение к другому нейрону.
Этапы развития пресинаптического торможения
Секреция тормозного медиатора (чаще ГАМК) аксоном тормозного нейрона
Взаимодействие медиатора с рецепторами мембраны возбуждающего аксона
2 сценария развития торможения
открытие анионных каналов → вхождение ионов Cl-
в клетку → нейтрализация ионов Na+, которые играют важную роль в развитии потенциала действия
(так как натрий +, а хлор -)
открытие потенциалзависимых К+-каналов →
К+ выходит из клетки во внеклеточную среду → гиперполяризация → мембранный потенциал становится более негативным → Ca2+ уже не может поступать в клетку, так как для него нужно, чтобы клетка была деполяризирована → «выключается» экзоцитоз медиатора
Клиническое значение
Пресинаптическое торможение развивается в афферентных нейронах задних рогов спинного мозга →
уменьшается поступление импульсов от болевых рецепторов.
Этот механизм присутствует у некоторых фармакологических препаратов, например, у морфина, так как он будет взаимодействовать с опиоидными рецепторами.

21
Механизм пресинаптического торможения
Воздействие возбуждающего нейрона
Развитие потенциала действия
Воздействие тормозного нейрона нейтрализует
потенциал действия
Нейромедиатор не высвобождается в синапс

22
1
2
3
4
5
6
7
8
9
10
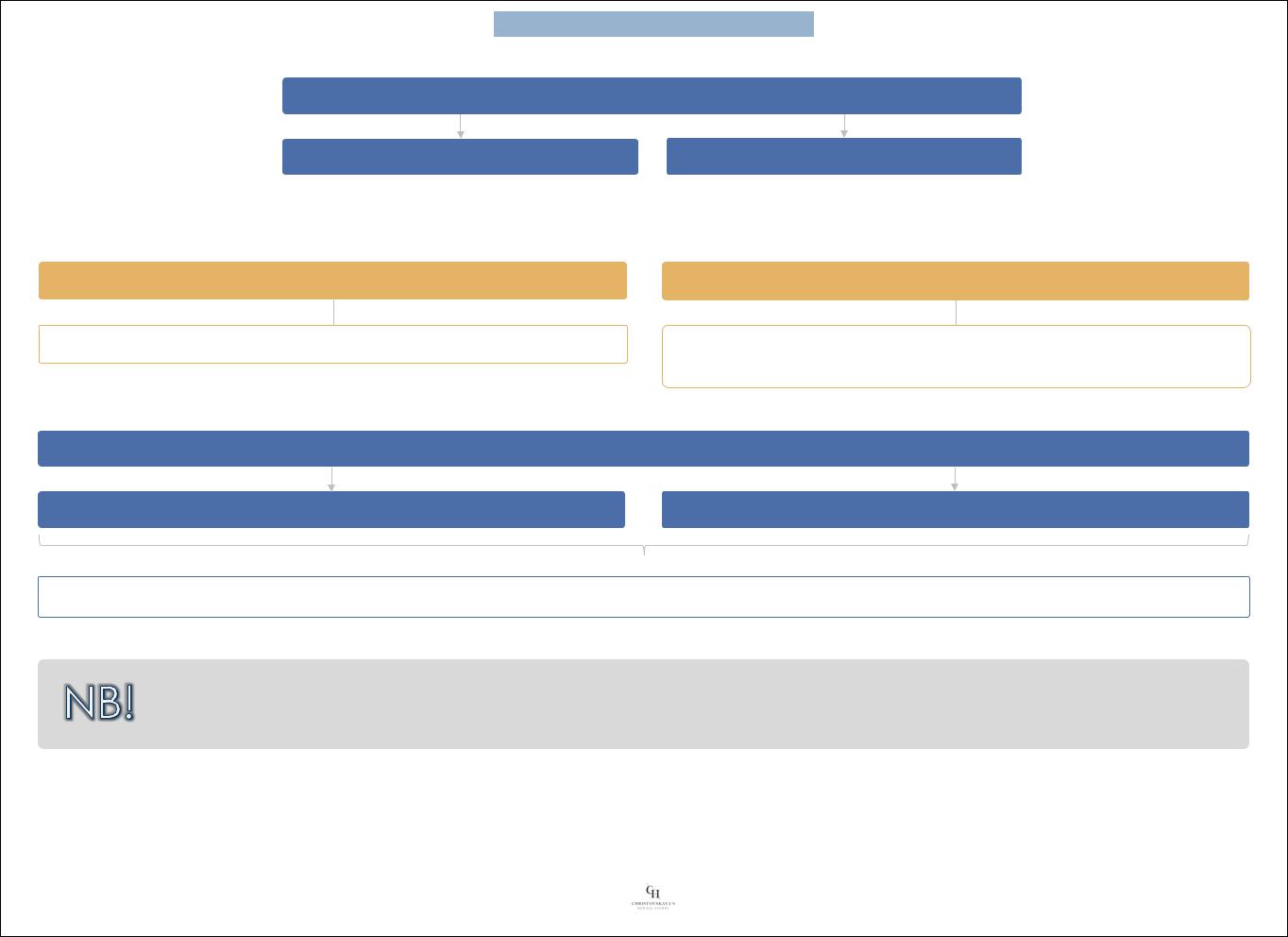
Места воздействия препарата в ЦНС:
Потенциальные цели препаратов
Потенциал действия в пресинаптическом волокне
Синтез нейромедиатора
Хранение нейромедиатора
Метаболизм нейромедиатора
Высвобождение нейромедиатора
Обратный захват нейромедиатора в пресинаптическое
окончание или захват в глиальную клетку
Разрушение нейромедиатора
Рецептор для нейромедиатора
Индуцированное рецептором увеличение
или уменьшение ионной проводимости
Отрицательная обратная связь
|
23 |
|
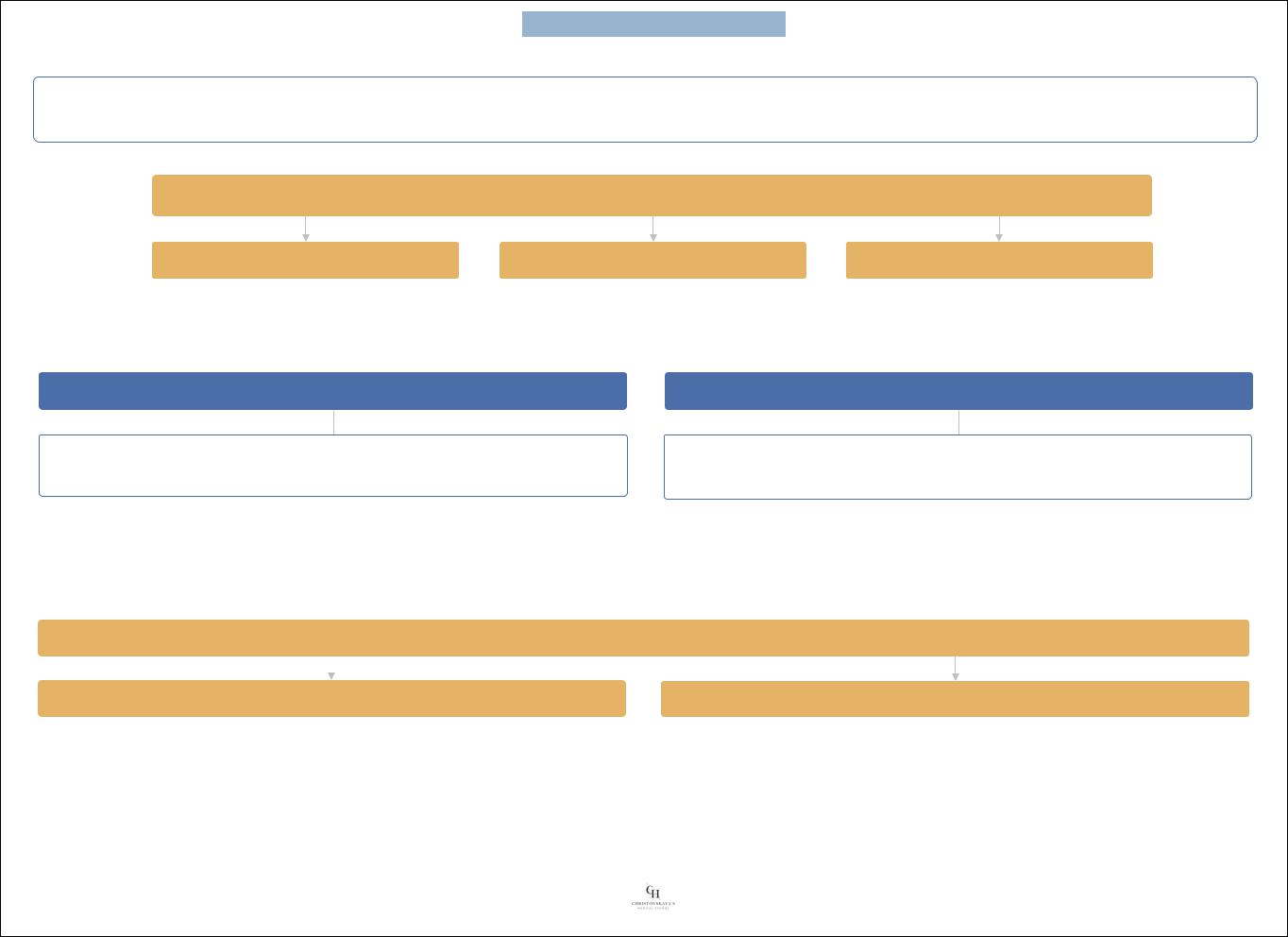
Клеточная организация мозга: |
|
|
Нейронные системы ЦНС |
|
|
иерархические |
неспецифические (диффузные) |
|
Иерархические нейронные системы |
||
Структура |
|
Функции |
чётко очерченные крупные миелинизированные волокна |
• |
сенсорное восприятие |
проводят потенциалы действия со скорость более 60 м/с |
• |
двигательный контроль |
|
|
|
Типы нейронов в ядрах и в коре головного мозга |
||
проекционные нейроны (интернейроны) |
|
нейроны локальный цепи |
два общих типа путей для этих нейронов образуют повторяющиеся пути обратной связи и пути прямой связи |
||
В сенсорных системах информация обрабатывается поочередно путём последовательных интеграций в каждом |
||
проекционном ядре на пути к коре головного мозга. |
|
|
Повреждение в любом звене выводит иерархическую нейронную систему из строя. |
||

24
Проекционные нейроны (интернейроны)
тела клеток большие, аксоны проецируются на большие расстояния, создают коллатерали и образовывают синапсы
с местными интернейронами
передают сигналы на большие расстояния
Характеристика: возбуждающие
Нейромедиатор: глутамат
Нейроны локальный цепи
тела клеток меньше, аксоны располагаются вблизи
от тела клетки. Образуют синапсы на теле проекционных нейронов, с дендритами тех же нейронов, между собой
особый класс нейронов локальной цепи в спинном мозге формирует
аксоаксонические синапсы на окончаниях сенсорных аксонов
Характеристика: тормозные Нейромедиаторы: ГАМК, глицин
Важно знать
Влияя на нейромедиаторы конкретных нейронов, можно воздействовать на общую возбудимость
центральной нервной системы.
Пример: пикротоксин – неконкурентный антагонист ГАМКА рецепторов. Его применение приводит к развитию генерализованных судорог.

25
Неспецифические (диффузные) нейронные системы
Структура
нейроны с тонкими немиелинизированными волокнами
проводят потенциалы действия медленно, со скоростью 0,5 м/с
Нейромедиаторы: моноамины, ацетилхолин
действуют преимущественно на метаботропные рецепторы → ответ реализуется долго
Отличия от иерархических систем
содержат ограниченное число нейронов, вырабатывающих
особые виды нейромедиаторов
Например: locus coeruleus (голубое пятно) – объединение
нейронов, вырабатывающих норадреналин. Ядро расположено в сером веществе моста (является частью ствола мозга), но нейроны проецируются по всему головному и спинному мозгу
Функции
вовлечены в регуляцию сна и бодрствование, внимания,
аппетита и эмоциональных состояний
Что это нам даёт?
Моноаминовые системы не могут передавать топографически специфические типы информации. Значит, обширные области ЦНС должны подвергаться воздействию препаратов одновременно и равномерно.
|
26 |
|
|
Нейромедиаторы в ЦНС: |
|
Механизм действия препаратов, влияющих на ЦНС, основывается на различии путей и действия нейромедиаторов → |
||
можно добиться относительной избирательности препаратов и снизить количество побочных эффектов. |
||
|
Аминокислотные медиаторы |
|
глутамат |
ГАМК |
глицин |
|
. |
|
|
Глутамат |
|
Функция |
|
Расположение |
осуществление возбуждающей |
|
широко распространённый нейромедиатор |
синаптической передачи |
|
в головном мозге |
|
|
накапливается в везикулах в пресинаптических окончаниях |
|
|
при помощи своего специфического транспортёра |
|
|
VGLUT (Vesicular Glutamate Transporter) |
|
Активирует рецепторы |
|
ионотропные |
|
метаботропные |
связаны с ионными каналами |
|
связаны с G-белком |

27
Путь глутамата
Глутамат высвобожается путем Са2+-зависимого экзоцитоза
Действует на постсинаптические рецепторы глутамата
Поступление глутамата в глию при помощи транспортёра →
преобразование в глютамин фермент: глютаминсинтетаза
Глютамин высвобождается из глии и поглощается нервным окончанием
Глютамин преобразуется в глутамат фермент: глютаминаза
Глутамин и глютамин – два варианта названия одной и той же аминокислоты.
Ионотропные рецепторы:
.
Механизм активации
Лиганд связывается с рецептором ионного канала
Изменяется проницаемость канала
Ионы проходят внутрь клетки

28
Виды ионотропных рецепторов:
AMPA
(рецептор α-амино-3-гидрокси-5- метил-4-изоксазолпропионовой кислоты)
•присутствуют во всех нейронах
•состоят из четырёх субъединиц
(GluA1–GluA4)
•рецепторы АМРА, содержащие субъединицу GluA2, проницаемы для Na+ и K+, но не для Ca2+
•рецепторы AMPA, лишённые субъединицы GluA2, проницаемы для Ca2+ (присутствуют на ингибирующих интернейронах)
KA
(каинатные рецепторы)
•распределены менее
равномерно, экспрессируются в
гиппокампе, мозжечке, спинном мозге
•состоят из субъединиц (GluK1–GluK5)
•проницаемы для Na+ и K+,
и в некоторых комбинациях субъединиц проницаемы для Ca2+
Активация этих рецепторов → открытие канала при потенциале покоя
NMDA
(N-метил-D-аспартат)
•присутствуют во всех нейронах ЦНС
•состоят из субъединиц
(GluN1 и одна/две субъединицы
GluN2 (GluN2A-GluN2D)
•обладают высокой проницаемостью для Ca2+, Na+, K+
•для открытия канала помимо глутамата необходим глицин
Требования для открытия канала
рецептора NMDA:
•глутамат и глицин должны связаться с рецептором
•мембрана должна быть деполяризована
Во время потенциала покоя просвет
блокируется внеклеточным Mg2+ →
активация рецептора не ведёт к открытию канала
При интенсивной активации соседних синапсов нейрон сильно деполяризован → Mg2+ вытесняется → канал открывается

29
Метаботропные рецепторы:
Связаны с G-белком. Реализуют свой ответ быстро (в течение секунд) через систему вторичных мессенджеров.
Имеют семь внутримембранных сегментов – sevenpass – змееподобные рецепторы.
Механизм активации:
Конец рецептора активирует гуанин-нуклеотид-связывающий белок (G-белок)
состоит из α, β, γ субъединицы: а-субъединица в норме связана
с гуанозиндифосфатом → делает G-белок неактивным
Белок меняет свою конформацию
Гуанозиндифосфат разъединяется с α-субъединицей
α-субъединица отделяется
Запуск других белков в сигнальном пути