

Шпоры к экзамену / chast_1
.docx
Остеогистогенез: классификация; 1) Эмбриональный остеогистогенез: прямой, непрямой. 2) Постэмбриональный остеогистогенез: регенерация. клетки костной ткани и их роль в остеогистогенезе; Остеобласты – молодые, функционально активные клетки. В зрелой кости местами их локализации являются: надкостница; эндост; каналы остеонов. Функции остеобластов: • остеобласты синтезируют и секретируют органический компонент межклеточного вещества (остеоид); Органические компоненты костной ткани: коллаген I типа, коллагены других типов, гликопротеины (остеонектин, остеокальцин), протеогликаны; • остеобласты осуществляют процесс минерализации костного матрикса – отложение кристаллов гидроксиапатита и других неорганических солей вдоль фибрилл коллагена - путѐм синтеза и секреции неколлагеновых белков, контролирующих процесс минерализации, в частности щелочной фосфатазы; • остеобласты участвуют в регуляции потока ионов Са и Р в костную ткань и обратно. • остеобласты синтезируют и выделяют различные ростовые факторы, в том числе и морфогенетические белки кости, определяющие превращение остеогенных клеток в остеобласты. Остеоцит – основной тип клеток зрелой костной ткани. Образуются из остеобластов, когда те в результате своей синтетической активности и минерализации оказываются окруженными со всех сторон минерализованным матриксом. Основная функция остеоцитов - поддержание нормального состояния остного матрикса и баланса кальция и фосфора в организме. Остеокласты имеют иное – гематогенное – происхождение. Остеокласты образуются из моноцитов крови путѐм их слияния с формированием многоядерных гигантских клеток (точнее, симпластов). Функции остеокластов: • остеокласты осуществляют разрушение (резорбцию) костной ткани; • поддержание минерального гомеостаза – в связи с высвобождением при разрушении матрикса большого количества минеральных веществ, поступающих в кровь. прямой остеогистогенез, его стадии и их характеристика; Прямой остеогенез характерен для развития грубоволокнистой костной ткани, образующей первоначально плоские кости черепа, ключицы, конечных фаланг пальцев. Он наблюдается очень рано, уже в первый месяц эмбриогенеза и включает три основные стадии: 1. Формирование остеогенного островка происходит путем скопления активно размножающихся клеток мезенхимы в участке развития будущей кости. 2. Дифференцировка клеток остеогенного островка и образование органического матрикса кости (остеоида). Клетки мезенхимы внутри остеогенного островка прекращают делиться и дифференцируются в остеобласты, вырабатывающие органический матрикс (остеоид). 3. Обызвествление остеоида обеспечивается остеобластами путем отложения кристаллов гидроксиапатита вдоль фибрилл коллагена и секреции матричных пузырьков. Замуровываясь в обызвествленном межклеточном веществе, остеобласты превращаются в остеоциты. непрямой остеогистогенез, его стадии и их характеристика; Непрямой остеогистогенез начинается на 2-м месяце внутриутробного развития и включает 5 стадий. 1. Формирование хрящевой модели (матрицы): осуществляется из скелетогенной мезенхимы. Образовавшаяся модель внешне сходна с будущей костью, образована гиалиновым хрящом, который снаружи покрыт надхрящницей и в течение некоторого времени увеличивается в размерах путём как аппозиционного, так и интерстициального роста. 2. Перихондральное окостенение диафиза: начинается в центральной части диафиза хрящевой модели с дифференцировки во внутреннем слое её надхрящницы остеобластов, которые секретируют костное межклеточное вещество. Образующиеся трабекулы из грубоволокнистой костной ткани формируют костную манжетку, которая располагается перихондрально. В последующем грубоволокнистая костная ткань в манжетке замещается на пластинчатую. 3. Эндохондральное окостенение диафиза: в центральной части диафиза в результате нарушения трофики начинаются дистрофия и последующая минерализация хрящевой ткани. Обызвествлённый хрящ разрушается остеокластами, формируются резорбционные каналы. Из накостницы врастают кровеносные сосуды, с которыми внутрь хрящевой модели проникают остеогенные клетки. 4. Эндохондральное окостенение эпифиза: начинается вскоре после рождения и осуществляется в результате процесса, сходного с ранее происходящим в диафизе. В обызвествлённый хрящ врастают кровеносные сосуды в окружении остеогенных клеток. 5. Формирование метэпифизарной хрящевой пластины роста. Между эпифизом и диафизом сохраняется дисковидная пластинка роста, образованная гиалиновой хрящевой тканью и имеющая четыре зоны: • зона неизмененного гиалинового хряща (граничит с эпифизом), • зона столбчатого хряща или монетных столбиков (широкая, содержит вертикальные изогенные группы хондроцитов, которые активно делятся, продуцируют межклеточное вещество и тем самым обеспечивают рост кости в длину), • зона пузырчатого (дистрофированного) хряща (состоит из «пузырчатых» хондроцитов), • зона минерализованного хряща (прилежит к эндохондральной кости диафиза), непрерывно разрушается и замещается разрастающейся эндохондральной костью. строение и функции метаэпифизарной хрящевой пластинки. Функция – рост кости в длину у детей и подростков.
|
Мышечные ткани классификация; В основу классификации мышечных тканей положены два принципа — морфофункциональный и гистогенетический. В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы. 1. Первая подгруппа — поперечнополосатые (исчерченные) мышечные ткани. В цитоплазме их элементов миозиновые филаменты постоянно полимеризованы, образуют с актиновыми нитями постоянно существующие миофибриллы. 2. Вторая подгруппа — гладкие (неисчерченные) мышечные ткани. Эти ткани характеризуются тем, что вне сокращения миозиновые филаменты деполимеризованы. В присутствии ионов кальция они полимеризуются и вступают во взаимодействие с филаментами актина. Образующиеся при этом миофибриллы не имеют поперечной исчерченности: при специальных окрасках они представлены равномерно окрашенными по всей длине (гладкими) нитями. В соответствии с гистогенетическим принципом в зависимости от источников развития (эмбриональных зачатков) мышечные ткани и мышечные элементы подразделяются на: • соматические (миотомные), • целомические (из миоэпикардиальной пластинки висцерального листка спланхнотома), • мезенхимные (из десмального зачатка в составе мезенхимы), • нейральные (из нервной трубки), • эпидермальные (из кожной эктодермы и из прехордальной пластинки). функции; Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Благодаря данной способности, мышечные ткани обеспечивают изменение положения в пространстве частей тела или тела в целом, а также изменение формы и объема отдельных органов. строение миосимпласта; Ядра миосимпласта - сравнительно светлые, с 1-2 ядрышками, диплоидные, овальные, уплощенные. Ориентированы длинной осью вдоль волокна. При резком сокращении волокон они могут укорачиваться, деформироваться и штопорообразно скручиваться. Содержание ядер несколько выше в красных волокнах по сравнению с белыми. Саркоплазма миосимпласта содержит все органеллы общего значения (за исключением центриолей) и некоторые специальные органеллы, а также включения. Сократительный аппарат мышечного волокна представлен миофибриллами - специальными органеллами, которые располагаются продольно в центральной части саркоплазмы и отделяются друг от друга рядами вытянутых митохондрий и цистерн саркоплазматической сети. Саркоплазматическая сеть - система уплощенных, вытянутых и анастомозирующих мембранных трубочек и мешочков, которая окружает каждый саркомер миофибриллы наподобие муфты. Саркоплазматическая сеть обладает выраженной способностью депонировать и выделять ионы кальция. Поперечные (Т-) трубочки представляют собой впячивания сарколеммы, отходящие от нее под прямым углом к оси волокна и расположенные у млекопитающих вблизи границы I- и А- дисков. Ветви соседних Т-трубочек опоясывают каждый саркомер и анастомозируют друг с другом. Митохондрии в миосимпласте располагаются в виде цепочек под сарколеммой и между миофибриллами. Они имеют вытянутую форму, содержат большое количество поперечно расположенных ламеллярных крист, характеризуются высокой активностью окислительно восстановительных ферментов. Их содержание и размеры больше в красных волокнах, чем в белых и увеличиваются при тренировке мышц. строение саркомера; Одно из следствий саркомерной структуры – это наличие у миофибрилл поперечной исчерченности. В каждой миофибрилле закономерно чередуются – светлые полосы, или I-диски (изотропные), шириной (в покое), – и темные полосы — А-диски (анизотропные. В частности, посередине каждого I-диска отчетливо видна Z-линия, или телофрагма. Саркомер — это участок миофибриллы между соседними Z-линиями. Следовательно, саркомер включает два полудиска I (прилегающие к соседним Z-линиям) и находящийся между ними диск А. В состав саркомера входят миофиламенты двух типов и опорные элементы, причем все эти компоненты имеют строго определенное положение. миосаттелитоциты и их роль в регенерации мышечных волокон. Миосателлитоциты - мелкие уплощенные клетки, располагающиеся в неглубоких вдавлениях сарколеммы миосимпластической части мышечного волокна и покрытые вместе с ней общей базальной мембраной. Ядро миосателлитоцита - плотное, относительно крупное (занимает почти всю клетку), с более высоким содержанием гетерохроматина, чем в ядрах миосимпласта, органеллы мелкие и немногочисленные. Эти клетки представляют собой камбиальные элементы скелетной мышечной ткани. Они активируются при повреждении мышечных волокон и обеспечивают их репаративную регенерацию. Слипаясь с симпластической частью волокна при усиленной нагрузке, миосателлитоциты участвуют в его гипертрофии.
|
Мышечные ткани строение сердечной мышечной ткани; Сердечная мышечная ткань образована клетками - кардиомиоцитами (сердечными миоцитами), связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих функциональных волокон. Различают два типа кардиомиоцитов: сократительные и проводящие. Сократительные кардиомиоциты: основная масса клеток; клетки покрыты сарколеммой, состоящей из плазмолеммы и базальной мембраны; 1-2 ядра в центре (полиплоидные); миофибриллы лежат по периферии клетки; их форма в желудочках цилиндрическая, в предсердиях - неправильная, часто отростчатая; клетки соединены конец в конец в области вставочных дисков, таким образом, сердечное волокно состоит из цепочек кардиомиоцитов, связанных вставочными дисками; связаны в единую трѐхмерную сеть благодаря наличию анастомозов, соединяющих соседние функциональные волокна; отличительной чертой предсердных миоцитов является слабое развитие Т-системы канальцев. В тех предсердных миоцитах, где нет Т-системы, на периферии клеток, под сарколеммой, располагаются многочисленные пиноцитозные пузырьки и кавеолы. Вставочный диск состоит из трѐх участков: - на вертикальных отрезках имеется: зона прикрепления миофибрилл, зона десмосом. - на горизонтальных отрезках: зона нексусов (коммуникация кардиомиоцитов, обеспечивает их синхронное сокращение). Особенности Т-системы сократительного кардиомиоцита (в отличие от скелетного мышечного волокна): характерны диады (состоят из двух компонентов: Т-трубочки и латеральной цистерны саркоплазматического ретикулума); Т-трубочки идут на уровне полоски Z. Проводящие кардиомиоциты обеспечивают ритмическое координированное сокращение отделов сердца; • Р-клетки (синусовые кардиомиоциты) – светло-окрашенные, мелкие, отростчатые, миофибрилл мало, ориентированы нерегулярно; локализация – сино-атриальный узел. Т-система отсутствует, но вдоль плазмолеммы находится много пиноцитозных пузырьков и кавеол, которые в 2 раза увеличивают мембранную поверхность клеток. Высокое содержание свободного кальция в цитоплазме этих клеток при слабом развитии саркоплазматической сети обусловливает способность клеток синусного узла генерировать импульсы. • Переходные клетки по строению и топографии занимают промежуточное положение между Р-клетками и сократительными кардиомиоцитами. Тонкие и вытянутые клетки. Функциональное значение этих клеток состоит в передаче возбуждения от Р-клеток к клеткам пучка и рабочему миокарду. • Клетки-волокна Пуркинье светлее, шире и короче сократительных кардиомиоцитов; на периферии цитоплазмы имеются немногочисленные неупорядоченные миофибрилл; обычно лежат пучками; Т-системы отсутствуют; нет типичных вставочных дисков; характерны включения гликогена, ферменты анаэробного гликолиза; преобладают в пучке Гиса и его ветвях.
|
Гладкая мышечная ткань строение; Источником развития гладкой мышечной ткани является в основном спланхнотомная мезенхима. Еѐ клетки мигрируют и окружают эпителиальные зачатки тех органов, в состав которых входит гладкая мышечная ткань. Гладкий миоцит – клетка веретеновидной формы, то есть их толщина максимальна в среднем участке, а к концам они конически сужаются. В цитоплазме у полюсов ядра концентрируются элементы секреторного аппарата: эндоплазматическая сеть – место синтеза белков межклеточного вещества, комплекс Гольджи, а также мелкие митохондрии. Для гладких миоцитов характерны многочисленные пузырьки, лежащие под цитолеммой, – кавеолы. Кавеолы и пузырьки редуцированного саркоплазматического ретикулума (СПР) депонируют ионы Са, необходимые для сокращения, и являются аналогом одновременно и саркоплазматического ретикулума и Т-трубочек в исчерченной мышечной ткани. Кавеолы и СПР содержат белки транспорта кальция. гистогенетическая классификация; Эндокринные гладкие миоциты (юкстагломерулярные, эпителиоидные, зернистые клетки) являются видоизмененными гладкими миоцитами, которые представляют собой основной компонент юкстагломерулярного аппарата почек. Они входят в состав стенки артериол почечного тельца и характеризуются редуцированным сократительным аппаратом при выраженном развитии синтетического аппарата. Продуцируемый этими клетками фермент ренин накапливается в их цитоплазме в виде покрытых мембраной гранул, содержимое которых выводится в кровь механизмом экзоцитоза. функции гладкой мышечной ткани; Сократительная. 1.Имеет свойства возбудимости и сократимости 2.Непроизвольное сокращение стенок внутренних половых органов. 3.Поднятие волос на коже 4.Подконтрольна вегетативной нервной системе регенерация гладкой мышечной ткани; Гладкая мышечная ткань способна к активной регенерации: • клеточная регенерация - за счѐт делений малодифференцированных клеток; • внутриклеточная регенерация – гипертрофия, восстановление органелл отличия в строении и функционировании поперечнополосатой и гладкой мышечных тканей. Гладкие мышцы характеризуются двумя особенностями. Во-первых, в отличие от скелетных мышц и миокарда они не имеют поперечной исчерченности (отсюда их название). Во-вторых, гладкие мышцы получают иннервацию не от соматического, а от вегетативного отдела нервной системы, поэтому лишены прямой произвольной регуляции. Гладкие мышцы пластичны, т.е. способны сохранять приданную растяжением длину без изменения напряжения. Скелетная мышца, наоборот, обладает малой пластичностью. Гладкомышечные волокна - веретенообразные клетки без поперечной исчерченности с одним ядром, способные к делению. Они содержат актиновые и миозиновые филаменты и сокращаются посредством механизма скользящих нитей. Скелетные мышцы в отличие от гладких способны совершать произвольные быстрые сокращения и производить этим значительную работу. Рабочим элементом мышцы является мышечное волокно. Мышечные волокна обладают тремя основными свойствами: возбудимостью — способностью отвечать на действия раздражителя генерацией потенциала действия; проводимостью — способностью проводить волну возбуждения вдоль всего волокна в обе стороны от точки раздражения; сократимостью — способностью сокращаться или изменять напряжение при возбуждении.
|
Нервная ткань: 1. Источники развития нервной ткани; Основной источник развития нервной ткани – нейроэктодерма. Из нее последовательно формируется нервная пластинка, нервный желобок и нервная трубка. При формировании нервной трубки часть клеток нервной пластинки не входит в ее состав и образует по бокам от нее нервный гребень или ганглиозную пластинку. В краниальном отделе по бокам нервной трубки формируется утолщение эктодермы – нейральные плакоды. Нейральные плакоды принимают участие в развитии 4 нервных узлов головы: V, VII, IX, X пар черепных нервов. Из нервной трубки в дальнейшем формируются нейроны и макроглия ЦНС. Ганглиозная пластинка дает начало нейронам и глиоцитам периферической нервной системы 2. Строение нейронов; Нейрон — основная клетка нервной ткани. Он имеет тело и отростки двух типов. В теле нейрона располагается ядро и органоиды, а по отросткам передаются нервные импульсы. Дендриты — это отростки, по которым нервные импульсы передаются к телу нейрона. Эти отростки сильно ветвятся. У нейрона может быть несколько дендритов. Аксон — это отросток, по которому импульсы передаются от тела клетки. 3. Морфологическая классификация нейронов; 1) мультиполярные – 1 аксон, много дендритов 2) униполярные – с одним аксоном (встречаются в эмбриональном развитии); 3) биполярные – с одним аксоном и одним дендритом (сетчатка глаза, спиральный ганглий); 4) псевдоуниполярные – дендрит и аксон отходят от нейрона в виде одного отростка, а затем разделяются (в спинномозговом ганглии). Это вариант биполярных нейронов. 4. Функциональная классификация нейронов 1) чувствительные (рецепторные, афферентные), их дендриты заканчиваются рецепторами (чувствительными нервными окончаниями). Воспринимают импульс 2) эффекторные (эфферентные), их аксоны заканчиваются двигательными или секреторными окончаниями. Передают импульс на ткани рабочих органов, побуждая их к действию. 3) ассоциативные (вставочными), осуществляют связь между нейронами. 5. Строение рефлекторной дуги. Рефлекторная дуга – система, состоящая из цепи нейронов, по которой проходит нервный импульс. В состав рефлекторной дуги входит один чувствительный (первый) и один эффекторный (последний) нейроны. Самая простая рефлекторная дуга состоит из цепи двух нейронов (афферентного и эфферентного). В состав сложных рефлекторных дуг входит более двух нейронов. Все нейроны рефлекторной дуги, кроме рецепторного и эффекторного, называются ассоциативными (вставочными).
|
Нервная ткань: 1. Классификация глии; Различают глию центральной и периферической нервной системы. Клетки глии центральной нервной системы делятся на макроглию (глиоциты) и микроглию. К макроглии относятся эпендимоциты, астроциты и олигодендроглиоциты. Микроглия представлена мелкими клетками, которые находятся в белом и сером веществе мозга К периферической нейроглии относятся нейролеммоциты и глиоциты ганглиев 2. Виды и функции астроцитов; Астроциты подразделяются на две группы: -протоплазматические (плазматические) – встречаются преимущественно в сером веществе ЦНС и имеют многочисленные разветвленные короткие отростки; -волокнистые (фиброзные) – располагаются в белом веществе ЦНС, имеют длинные отростки. Функции астроцитов: -опорная; -барьерно-защитная (участие в образовании гематоэнцефалического барьера; имеют выраженную способность к фагоцитозу); -транспортная; -разграничительная; -регуляторная и метаболическая (участие в обмене нейромедиаторов) 3. Олигодендроглия и её функции; Олигодендроглия – мелкие клетки с короткими отростками, окружают тела нейронов, входят в состав нервных волокон и нервных окончаний. Характеризуются темным ядром, плотной цитоплазмой. Функции олигодендроглии: -барьерно-защитная; -выработка миелина – образование глиальной оболочки нервных волокон; -регуляция метаболизма нейронов. 4. Виды эпендимоцитов и их функции; Эпендимоциты: собственно эпендимоциты, хороидные эпендимоциты и танициты Функции эпендимной глии: - опорная; - образование нейро-ликворного и гемато-ликворного барьеров; - ультрафильтрация компонентов СМЖ. 5. Источник происхождения и функции микроглии. Имеют мезенхимное происхождение. Клетки небольших размеров с уплотненной цитоплазмой и тонкими отростками, с большим содержанием лизосом. В ядрах преобладает гетерохроматин. Функции микроглии: клетки активно передвигаются по нервной ткани и проявляют фагоцитарную активность, поглощают гибнущие нейроны и нервные волокна. В зависимости от состояния, встречаются 3 вида микроглии: - в развивающемся мозгу – амёбоидная микроглия (способна к перемещению и активно фагоцитирует фрагменты разрушающихся клеток) - в сформированном мозгу – покоящаяся (ветвистая) микроглия (слабая фагоцитарная активность), - в травмированном мозгу – реактивная микроглия (вновь – высокая фагоцитарная активность).
|
Нервные волокна: 1. Строение миелиновых нервных волокон
1
2 1 — ядро леммоцита (шванновской клетки): располагается в центре волокна; 2 — осевые цилиндры (отростки нейронов): 10—20 осевых цилиндров погружено по периферии волокна в цитоплазму леммоцита. Над каждым цилиндром плазмолемма леммоцита смыкается - так. что образуется "брыжейка", или 3 — мезаксон; 4 — базальная мембрана вокруг нервного волокна.
3. механизм и скорость проведения нервного импульса; В безмиелиновом волокне волна идет по всей аксолемме, не прерываясь, а в миелиновом возникает только в области перехвата, проведение возбуждения прыжками. Между перехватами идет электрический ток, скорость которого выше, чем прохождение волны деполяризации по аксолемме. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1-2 м/с, тогда как толстые миелиновые – со скоростью 5-12 м/с. 4. регенерация нервных волокон после повреждения; Регенерация нервных волокон в ЦНС отсутствует: хотя нейроны ЦНС обладают способностью к восстановлению своих отростков, этого не происходит, по-видимому, вследствие неблагоприятного влияния микроокружения. После повреждения нейрона микроглия, астроциты и гематогенные макрофаги фагоцитируют детрит в участке разрушенного волокна, на его месте пролиферирующие астроциты образуют плотный глиальный рубец 5. строение рефлекторной дуги. Рефлекторная дуга – система, состоящая из цепи нейронов, по которой проходит нервный импульс. В состав рефлекторной дуги входит один чувствительный (первый) и один эффекторный (последний) нейроны. Самая простая рефлекторная дуга состоит из цепи двух нейронов (афферентного и эфферентного). В состав сложных рефлекторных дуг входит более двух нейронов. Все нейроны рефлекторной дуги, кроме рецепторного и эффекторного, называются ассоциативными (вставочными).
|
Нервные окончания: 1. определение и виды нервных окончаний Нервные окончания — это концевые аппараты нервных волокон, различные по функциональному значению. 1) концевые аппараты, образующие межнейрональные синапсы 2) эффекторные окончания– передают нервный импульс на ткани рабочего органа. 3) рецепторные (аффекторные, или чувствительные) - окончания дендритов рецепторных (чувствительных) нейронов. 2. классификация синапсов Классификация: 1)по локализации: - центральные (ГМ,СМ) - периферические (нервно-мышечный) 2) по механизму передачи сигнала: - химические; - электрические - смешанные. 2) по выделяемому нейромедиатору: - адренергические – нейромедиатор норадреналин; - холинергические – нейромедиатор ацетилхолин; - дофаминергические – нейромедиатор дофамин; - серотонинергические – нейромедиатор серотонин; - ГАМК-ергические – нейромедиатор гамма-аминомасляная кислота (ГАМК) 3) по влиянию: - возбуждающие; - тормозные. 4) по местоположению: - нервно-мышечные; - нейро-нейрональные: а) аксо-соматические; б) аксо-аксональные; в) аксо-дендрические; г) дендро-соматические. Структурно-функциональная характеристика синапсов. 1)пресинаптическое окончание: а) пресинаптическая мембрана б) пузырьки/везикулы – заполненные медиаторы в) митохондрии 2) синаптическая щель – содержит межклеточную жидкость и мукополисахаридное плотное в-во в виде полосок, мостиков, которое обеспечивает связь между пресинаптической и постсинаптической мембранами и может содержать ферменты 3) постсинаптическая мембрана- утолщенная часть клеточной мембраны иннервируемой клетками, содержащая белковые рецепторы, имеющие ионные каналы и способные связать молекулы медиатора 3. Строение химического синапса Химические синапсы передают нервные импульсы только в одном направлении и с задержкой. Химические синапсы имеют все три компонента. В пресинаптической части находятся синаптические пузырьки с медиатором, митохондрии, аЭПС, нейротрубочки и нейрофиламенты. Строение синаптических пузырьков зависит от содержащегося в них медиатора: - мелкие и электронно-прозрачные пузырьки характерны для холинергических синапсов; - крупные, с электронноплотной частью в центре – для адренегрических синапсов; - крупные, с плотной сердцевиной и светлым ободком - для пептидергических синапсов. Ширина синаптической щели около 30 нм. В ней содержатся элементы гликокаликса. Постсинаптическая часть содержит постсинаптическое уплотнение. Постсинаптическая мембрана содержит интегральные белки - синаптические рецепторы, связывающиеся с нейромедиатором. 4. Морфологическая классификация рецепторов; свободные; несвободные (инкапсулированные: пластинчатые тельца Фатера-Пачини, осязательные тельца Мейснера, концевые колбы Краузе, сухожильные органы Гольджи; неинкапсулированные); 5. Простая и сложная рефлекторные дуги. Самая простая рефлекторная дуга состоит из цепи двух нейронов (афферентного и эфферентного). В состав сложных рефлекторных дуг входит более двух нейронов.
|
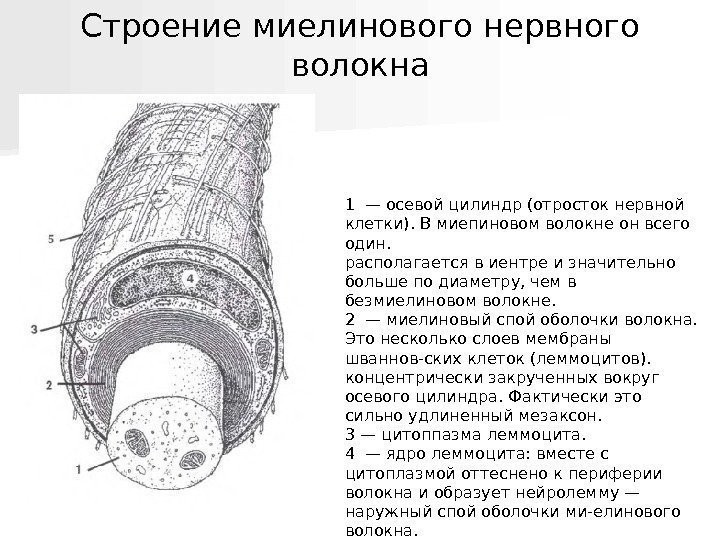 - осевой цилиндр (отросток нервной
клетки). В миелиновом волокне он всего
один.
располагается в центре и
значительно больше по диаметру, чем
в безмиелиновом волокне.
2 - миелиновый
спой оболочки волокна.
Это несколько
слоев мембраны шванновских клеток
(леммоцитов). концентрически закрученных
вокруг осевого цилиндра. Фактически
это сильно удлиненный мезаксон.
3
— цитоплазма леммоцита.
4 - ядро
леммоцита: вместе с цитоплазмой
оттеснено к периферии волокна и
образует нейролемму - наружный спой
оболочки миелинового волокна.
- осевой цилиндр (отросток нервной
клетки). В миелиновом волокне он всего
один.
располагается в центре и
значительно больше по диаметру, чем
в безмиелиновом волокне.
2 - миелиновый
спой оболочки волокна.
Это несколько
слоев мембраны шванновских клеток
(леммоцитов). концентрически закрученных
вокруг осевого цилиндра. Фактически
это сильно удлиненный мезаксон.
3
— цитоплазма леммоцита.
4 - ядро
леммоцита: вместе с цитоплазмой
оттеснено к периферии волокна и
образует нейролемму - наружный спой
оболочки миелинового волокна.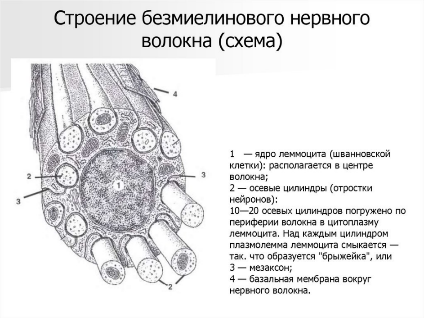 .
строение безмиелиновых нервных волокон
.
строение безмиелиновых нервных волокон