

Инициация
Начинается с активации промотора с помощью белка ТАТА-фактора, который взаимодействует со специфической последовательностью нуклеотидов промотора -ТАТААА- (ТАТА-бокс).
Присоединение ТАТА-фактора облегчает взаимодействие промотора с ДНК-зависимой-РНК- полимеразой.
Это вызывает изменение конформации РНК-полимеразы то и обеспечивают раскручивание См примерно одного витка спирали ДНК и локальное плавление (разрушение водородных связей), т.о. образуется транскрипционная вилка.
Элонгация
На участке примерно в 40 пар нуклеотидов РНК- полимераза по принципу комплементарности образует фосфоэфирные связи между нуклеотидами, участвуя в синтезе полинуклеотидной цепи молекулы мРНК от 5'-к З' концу.
Скорость элонгации - примерно 50 нуклеотидов в секунду. Т.о. образуется полная комплементарная копия транскриптона от промотора до терминатора, т.е образуется первичная структура мРНК.
Терминация
Транскрипция завершается после достижения РНК- полимеразой сайта терминации, который имеет особую структуру в виде палиндрома.
Палиндром — это слова или фраза, которые одинаково читаются слева направо. (участок связанных комплиментарных нуклеотидов одной цепи РНК или ДНК, в котором последовательность нуклеотидов совпадает с комплиментарной ей последовательностью при чтении в направлении от 5'-конца к 3'-концу)
В результате синтеза палиндрома формируется структура матричной РНК, которая называется «шпилькой» и РНК- полимераза отсоединяется от цепи ДНК. Окончанию терминации способствуют белковые факторы терминации.
У прокариот образованная мРНК сразу участвует в трансляции белка и эти два процесса происходят практически одновременно. У эукариот продуктом транскрипции является прематричная РНК, которая в дальнейшем подвергается процессу созревания или процессингу.
Процессинг мРНК у эукариот.
В результате транскрипции образуется первичный транскрипт - строго комплементарная матрице пре-мРНК. Для повышения устойчивости молекулы, на стадии 30 нуклеотидов в ядре происходит:
Кэпирование - присоединение к 5' концу пре-мРНК метилированного гуанозина, с помощью фосфодиэфирной связи. Кэпирование защищает молекулу мРНК от эндонуклеаз и способствует распознаванию первого кодона при синтезе белка.
30.09.2024 Продолжение
Полиаденилирование- присоединение к 3 концу пре-мРНК ферментом полимеразой от 30 до 300 адениловых нуклеотидов. Полиаденилирование облегчает выход мРНК из ядра и заменят её гидролиз в цитоплазме.
Сплайсинг. Последовательности нуклеотидов, присутствующие в ДНК, но не входящие в состав зрелой мРНК, были названы некодирующими или интроны, а последовательности присутствующие в мРНК- кодирующие или экзоны. Последовательность интронов «вырезаются» из первичного транскрипта, концы экзонов соединяются друг с другом.
Такую модификацию РНК называют «сплайсинг», происходит в ядре, в цитоплазму поступает уже «зрелая» мРНК.
Активация аминокислот – второй подготовительный процесс.
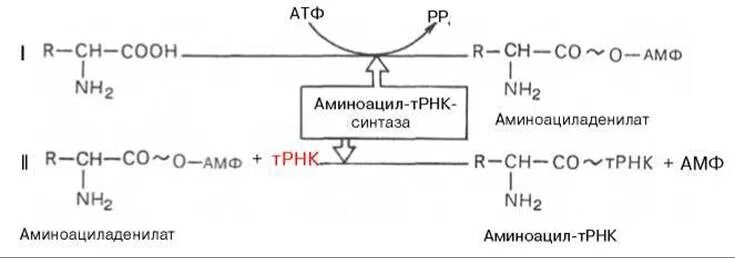
Взаимодействие АК с АТФ и специфическим ферментом – образуется активированный комплекс
Перенос ферментом АК на свою тРНК
Таким образом в рибосому поступает информация (в виде мРНК) и активированный комплекс, несущий вещество (АК) и энергию (в виде макроэргической связи)
Воспроизведение идёт с помощью генетического кода – система записи информации о последовательности расположения аминокислот в белках с помощью последовательности нуклеотидов мРНК. (в таблице для мРНК)
Свойства генетического кода
Триплетность- каждая АК кодируется 3 расположенными рядом нуклеотидами (кодонами)
Выраженность или избыточность – 64 триплета кодируют 20 АК
Однозначность- - каждый триплет кодирует только одну аминокислоту
Полярность – информация считывается в одном направлении, поэтому есть кодоны, с которых начинается считывание, и стоп-кодоны
Неперекрываемость – каждый нуклеотид может входить в состав только одного триплета
Универсальность – код един для всех организмов
Рибосомы.
Когда синтез белка непосредственно не идет, рибосомы находятся в цитоплазме в диссоциированном виде. В момент начала синтеза белка мРНК обратимо присоединяется к поверхности малой субъединицы рибосомы в присутствии ионов магния. При этом два первых транслируемых триплета (кодона) обращены к большой субъединице рибосомы.
Первый кодон связывает молекулу транспортной РНК, несущую первую аминокислоту, дальше к комплексу: малая субъединица рибосомы - мРНК- тРНК-аминокислота присоединяется большая субъединица рибосомы
Первым кодоном мРНК всегда является АУГ; он всегда присоединяет к себе антикодон тРНК, которая несет аминокислоту -формил-метионин -эта аминокислота не войдёт в состав белка, её функция – соединить две субъединицы рибосомы.
Механизм трансляции
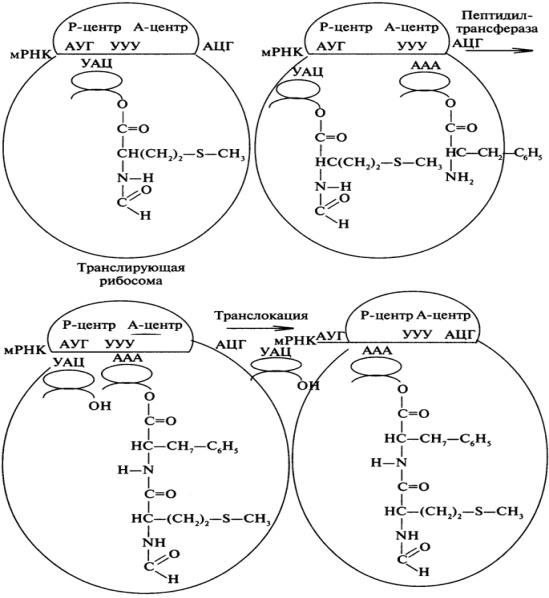
Инициация – связывание антикодонов тРНК с кодоном мРНК по принципу комплиментарности в аминоацильном участке рибосомы. Перемещение мРНК на 1 кодон в пептидильный участок рибосомы. Поступление в аминоацильный участок новой тРНК со своей АК.
Элонгация – 1 шаг – транспептизация – удлинение цепи – переброс растущего пептида на аминогруппу вновь вошедшей аминокислоты. Образование пептидной связи. 2 шаг- трансмембранное перемещение рибосомы относительно мРНК на один кодон.
Терминация – узнавание рибосомой стоп-кодона и отсоединение новосинтезированного белка.
Посттрансляционная модификация белка — формирование вторичной, третичной и четвертичной структуры белка при участии ферментов и с затратой энергии.
Модификации боковых цепей аминокислот (в ЭПР или КГ)
Гликозилирование
Гидроксилирование
Ацетилирование
Метилирование
Карбоксилирование
Фосфорилирование
В клетки синтез одного не очень сложного белка завершается через одну-две минуты.