

27.09.2024 Дыхание микроорганизмов
Классификация бактерий по типу дыхания:
облигатные аэробы
микроаэрофилы
факультативные анаэробы
облигатные анаэробы
Облигатные (строгие) аэробы развиваются при наличии в атмосфере 20% кислорода (микобактерии туберкулеза) содержат ферменты, с помощью которых осуществляется перенос водорода от окисляемого субстрата к кислороду воздуха.
Микроаэрофилы нуждаются в значительно меньшем количестве кислорода, и его высокая концентрация хотя и не убивает бактерии, но задерживает их рост (актиномицеты, бруцеллы, лептоспиры)
Факультативные анаэробы могут размножаться как в присутствии, так и в отсутствие кислорода (большинство патогенных и сапрофитных микробов — возбудители брюшного тифа, паратифов, кишечная палочка).
Облигатные анаэробы — бактерии, для которых наличие молекулярного кислорода является губительным (клостридии столбняка, ботулизма)
Аэробное дыхание является самой совершенной формой окислительного процесса и наиболее эффективным способом получения энергии
Г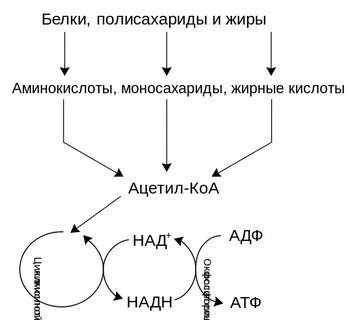 лавное
преимущество аэробного дыхания состоит
в том, что энергия окисляемого вещества
- субстрата, на котором микроорганизм
растет, используется наиболее полно.
Поэтому в процессе дыхания перерабатывается
гораздо меньше субстрата для получения
определенного количества энергии, чем,
например, при брожениях.
лавное
преимущество аэробного дыхания состоит
в том, что энергия окисляемого вещества
- субстрата, на котором микроорганизм
растет, используется наиболее полно.
Поэтому в процессе дыхания перерабатывается
гораздо меньше субстрата для получения
определенного количества энергии, чем,
например, при брожениях.
Процесс дыхания заключается в том, что углеводы (или белки, жиры и другие запасные вещества клетки) разлагаются, окисляясь кислородом воздуха, до углекислого газа и воды. Выделяющаяся при этом энергия расходуется на поддержание жизнедеятельности организмов, рост и размножение. Бактерии вследствие ничтожно малых размеров своего тела не могут накапливать значительного количества запасных веществ. Поэтому они используют в основном питательные соединения среды.
Упрощённая схема трёхстадийного распада молекул питательных веществ
Р
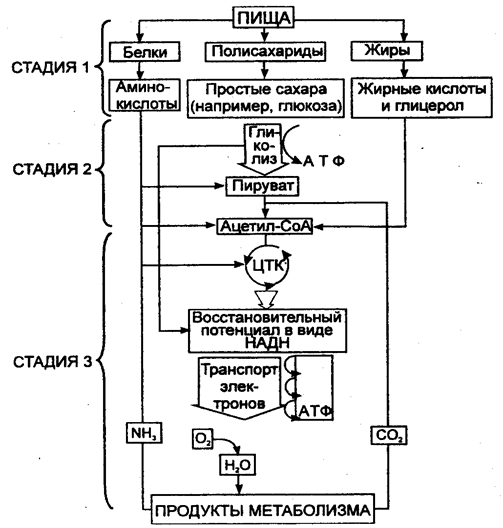 асщепление
макромолекул на простые субъединицы
асщепление
макромолекул на простые субъединицыРасщепление простых молекул до ацетила-Со А сопровождавшееся выделением АТФ и НАДН
Полное окисление ацетил-Со А до воды и углекислого газа, что сопровождается выделением АТФ
В общем виде дыхание можно представить следующим уравнением:
С6Н12О6 + 6О2 = 6СО2 + 6Н2О + 2875 КДЖ
За этой простой формулой скрывается сложная цепь химических реакций, каждая из которых катализируется специфическим ферментом.
Ферментативные реакции происходящие в процессе дыхания, в настоящее время хорошо изучены. Схема реакций оказалась универсальной, т. е. в принципе одинаковой у животных, растений и многих микроорганизмов, в том числе бактерий.
С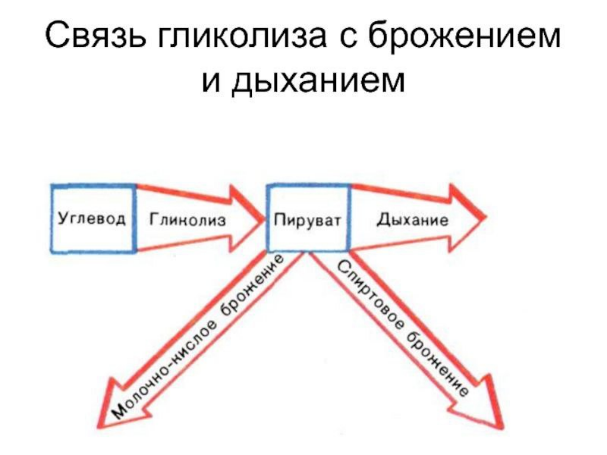 начала
происходит образование фосфорных эфиров
глюкозы- монофосфата, затем дифосфата
Фосфорная кислота переносится ферментами
(трансферазами) с аденозинтрифосфорной
кислоты (АТФ)- вещества, имеющего три
остатка фосфорной кислоты, соединенных
макроэргическими связями На присоединение
фосфорной кислоты тратится 3,4.Л0'Дж
опергии на І грамм-молекулу Поэтому
образовывается связь называется
макроэргической, Биологический смысл
первых реакций фосфорилирования
заключается в активированной глюкозы
- присоединение фосфора к глюкозе делает
ее более реакционноспособной, лабильной,
определяет возможность дальнейшего
расщепления глюкозы.
начала
происходит образование фосфорных эфиров
глюкозы- монофосфата, затем дифосфата
Фосфорная кислота переносится ферментами
(трансферазами) с аденозинтрифосфорной
кислоты (АТФ)- вещества, имеющего три
остатка фосфорной кислоты, соединенных
макроэргическими связями На присоединение
фосфорной кислоты тратится 3,4.Л0'Дж
опергии на І грамм-молекулу Поэтому
образовывается связь называется
макроэргической, Биологический смысл
первых реакций фосфорилирования
заключается в активированной глюкозы
- присоединение фосфора к глюкозе делает
ее более реакционноспособной, лабильной,
определяет возможность дальнейшего
расщепления глюкозы.
Первая - анаэробная-стадия процесса дыхания носит название гликолитического пути или пути Эмбдена - Мейергофа- Парнаса. Для осуществления этих реакций кислород не требуется. Образовавшаяся пировиноградная кислота (СНЗСОСООН) является интереснейшим и очень важным соединением. Пути расщепления глюкозы в процессе дыхания и многих брожений вплоть ДО образования пировиноградной кислоты идут совершенно одинаково, что впервые было установлено русским биохимиком С. П. Костычевым. Пировиноградная кислота является тем центральным пунктом, от которого расходятся пути дыхания и брожений, откуда начинается специфическая для данного процесса цепь ферментативных превращений - специфическая цепь химических реакций
Дыхание является процессом, дающим энергию, однако его биологическое значение этим не ограничивается. В результате химических реакций, сопровождающих дыхание, образуется большое количество промежуточных соединений. Из этих соединений, имеющих различное количество углеродных атомов, могут синтезироваться самые разнообразные вещества клетки: аминокислоты, жирные кислоты, жиры, белки, витамины. Поэтому обмен углеводов определяет остальные обмены веществ (белков, жиров). В этом его огромное значение.
Цикл Кребса — это цепь химических реакций, которая называется циклом потому, что продолжается непрерывно. цикл трикарбоновых кислот — это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения множества метаболических путей, промежуточный лап между гликолизом и электрон-транспортной цепью. Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в холе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения как аминокислоты, углеводы, жирные кислоты и др. Для осуществления всего вышеизложенного необходимо наличие более ста различных элементов, в том числе витаминов. При отсутствии или недостатке хотя бы одного из них цикл будет недостаточно эффективным, что приведёт к нарушению метаболизма.
Х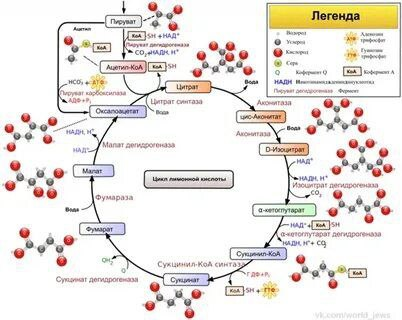 отя
в ходе цикла трикарбоновых кислот
непосредственно образуется лишь одна
молекула АТФ на оборот (при превращении
сукцинил-КоА в сукцинат) четыре
окислительные реакции цикла обеспечивают
дыхательную цепь значительным числом
электронов поставляемых НАДН и ФАДН2,
и тем самым обеспечивают образование
значительного количества АТФ в ходе
окислительного фосфорилирования
отя
в ходе цикла трикарбоновых кислот
непосредственно образуется лишь одна
молекула АТФ на оборот (при превращении
сукцинил-КоА в сукцинат) четыре
окислительные реакции цикла обеспечивают
дыхательную цепь значительным числом
электронов поставляемых НАДН и ФАДН2,
и тем самым обеспечивают образование
значительного количества АТФ в ходе
окислительного фосфорилирования
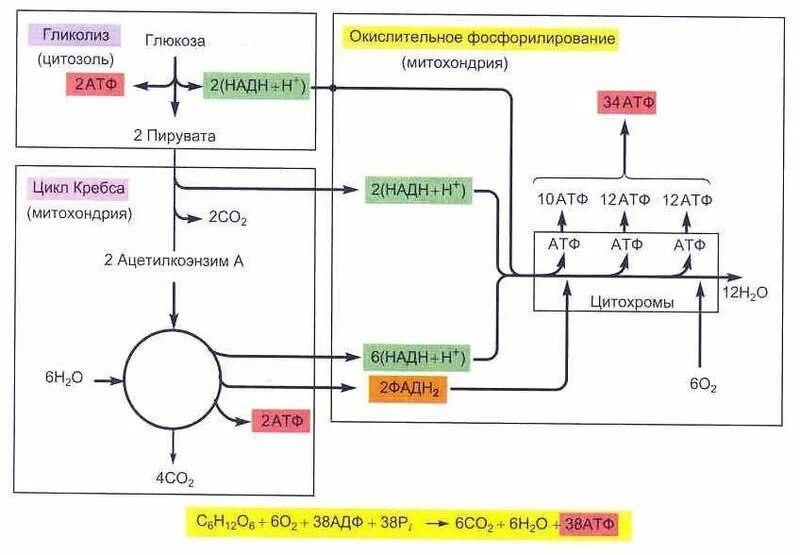
В ходе гликолиза из одной молекулы глюкозы образуется две молекулы пирувата, 2 АТФ и 2 НАДН. В ходе окислительного фосфорилирования переход двух электронов с НАДН на 02 обеспечивает образование 2,5 АТФ, а переход двух электронов с ФАДН2 на 02 даёт1,5 АТФ. Когда обе молекулы пирувата окисляются до 6 СО2пируватдегидрогеназным комплексом и в ходе цикла трикарбоновых кислот, а электроны переносятся на 02 в ходе окислительного фосфорилирования, то суммарный выход АТФ составляет 32 молекулы на молекулу гюкозы.
Дыхательная цепь переноса электронов, также электрон-транспортная цепь.
(сокр. ЭТЦ) — система трансмембранных белков и переносчиков электронов, необходимых для поддержания энергетического баланса ЭТЦ поддерживает баланс за счёт переноса электронов п протонов из НАД·Н и ФАДН2 в акцептор электронов. В случае аэробного дыхания акцептором может быть молекулярный кислород (02). В случае анаэробного дыхания акцептором могут быть NO3-, NO2-, Fe3+, фумарат, диметилсульфоксид, сера, SO4 2-, CO2 и т.п.
ЭТЦ у прокариот локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики электронов расположены в порядке уменьшения сродства к электрону, то есть по своему окислительно-восстановительному потенциалу, где у акцептора самое сильное сродство к Электрону. Поэтому транспорт электрона на всём протяжении цепи протекает самопроизвольно с выделением энергии. Энергия, выделяющаяся при переносе электронов, выделятся в межмембранное пространство ступенчато, в виде протона (Н+), выделясь из каждого комплекса. Протоны из межмембранного пространства попадают в протонную помпу. Там протоны создают протонный потенциал. Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ Сопряжённая работа ЭТЦ и АТФ-спнтазы носит название окислительного фосфорилирования.
Анаэробное дыхание цепь анаэробных окислительно- восстановительных реакций, которые сводятся к окислению органического или неорганического субстрата с использованием в качестве конечного акцептора электронов не молекулярного кислорода, а других неорганических веществ (нитрата - NО3-, нитрита- NO2-, сульфата-SO4 2-, сульфита-SO3 2-,СO2 др.), а также органических веществ (фумарата и др.) Молекулы АТФ в процессе анаэробного дыхания образуются в основном в электрон-транспортной цепи, T. е. В результате реакций мембранного фосфорилирования, но в менышем количестве, чем при аэробном дыхании.
А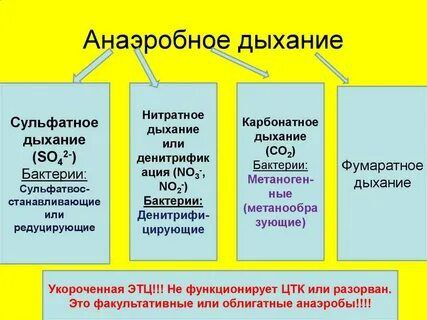 наэробное
дыхание
наэробное
дыхание
При анаэробном дыхании конечным акцептором электронов в электрон-транспортной цепи являются неорганические или органические соединения. Например, если конечным акцептором электронов является SO 2-, то процесс называют сульфатным дыханием, а бактерии — сульфатвосстанавливающими или сульфатредуцирующими В том случае, если конечным акцептором электронов служит NО3 - или NО2-, то процесс называется нитратным дыханием или денитрификацией, а бактерии, осуществляющие этот процесс, — денитрифицирующими В качестве конечного акцептора электронов может выступать СО2. Процесс соответственно называют карбонатным дыханием, а бактерии - метаногенными (метанобразующими) Одним из немногих примеров, когда конечным акцептором служит органическое вещество, является фумаратное дыхание
Основные особенности бактерий, способных к анаэробному дыханию
Имеют укороченные электрон-транспортные, или дыхательные, цепи ,т. е. они не содержат всех переносчиков, характерных для дыхательных цепей, функционирующих в аэробных условиях
В дыхательных цепях анаэробов цитохромоксидаза заменена соответствующими редуктазами
У строгих анаэробов не функционирует цикл Кребса, выполняет только биосинтетические функции
Основное количество молекул АТФ при анаэробном дыхании синтезируется в процессе мембранного фосфорилирования
По отношению к молекулярному кислороду бактерии, осуществляющие анаэробное дыхание, являются факультативными пли облигатным анаэробами. К облигатным анаэробам относятся сульфатвосстанавливающее и метаногенные бактерии. К факультативным анаэробам = денитрифицирующие бактерии и бактерии, осуществляющие фумаратное дыхание. Факультативные анаэробы могут переключать свой энергетический метаболизм с аэробного дыхания в присутствии в среде молекулярного кислорода на анаэробное дыхание в отсутствии молекулярного кислорода
Выход АТФ при анаэробном дыхании меньше, чем при аэробном, но больше, чем при брожении
Брожение
метаболический процесс, при котором регенерируется АТФ, а продукты расщепления органического субстрата могут служить одновременно и донорами, и акцепторами водорода. Брожение —это анаэробный (происходящий без участия кислорода) метаболический распад молекул питательных веществ, например глюкозы. Больтинство типов брожения осуществляют микроорганизмы облигатные или факультативные анаэробы
Любое брожение проходит в две стадии:
первая (окисление) включает превращение глюкозы в пировиноградную кислоту.
вторая (восстановление) — использование атомов водорода для восстановления пировиноградной кислоты
Брожение не высвобождает всю имеющуюся в молекуле энергию, поэтому промежуточные продукты брожения могут использоваться в ходе клеточного дыхания
Б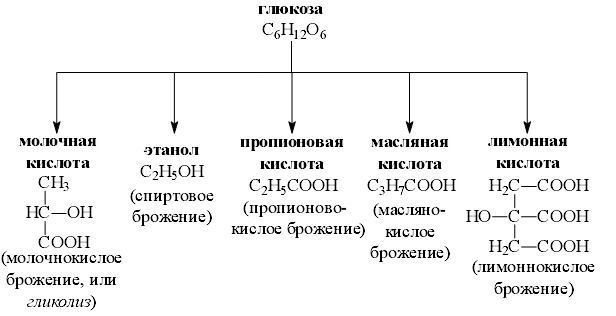 рожение:
рожение:
4. муравьинокислое (смешанное брожение — характерным, хотя не главным, продуктом является муравьиная кислота НСООН);
5. маслянокислое (образуется масляная кислота СЗН7СООН);
6. уксуснокислое (окисление этилового спирта в уксусную кислоту СНЗСООН).
Ферменты
Оксидоредуктазы= катализируют окислительно-восстановительные реакции.
Трансферазы осуществляют реакции переноса груши атомов.
Гидролазы= осуществляют гидролитическое расщепление различных соединений ,
Лиазы- катализируют реакции отщепления от субстрата химической группы негидролитическим путем.
Лигазы или синтетазы =обеспечивают соединение двух молекул, сопряженное с расщеплением пирофосфатной связи в молекуле АТФ
Изомеразы = определяют пространственное расположение групп элементов
По типу выработке: конститутивные- клеткой постоянно, нужны для обмена в-в; индуцибельные- при наличии индуктора, репрессивные
По локализации в клетке: эндоферменты, экзоферменты (гиалуронидаза, нейраминидаза)
Микробиологическая (рабочая) классификация ферментов:
1. Сахаролитические
2. Протеолитические
3. Липолитические
4. Окислительно-восстановительные (каталаза, цитохромоксидаза)
5. Аутолитические
6. Ферменты патогенности (вирулентности)