

Лекции Хомича Цитология
.pdfможливості формування апарату білкового синтезу. Цитоплазма (овоплазма) займає великий об’єм. У ній є вільні рибосоми, ендоплазматична сітка, комплекс Гольджі, мітохондрії, лізосоми і жовток. Жовток складається з білків, вуглеводів і ліпідів. Накопичуючись у цитоплазмі, він зміщує ядро на периферію, у зв’язку з чим яйцеклітина набуває полярності. Жовток є `трофічним матеріалом для зародка. Його кількість у яйцеклітині пов’язана з типом ембріонального розвитку тварин – внутрішнім або зовнішнім, а також від виду зовнішнього типу – прямого чи непрямого.
Залежно від кількості жовтка яйцеклітини поділяють на оліголецитальні, полілецитальні й мезолецитальні.
Оліголецитальні яйцеклітини містять мало жовтка. Вони властиві тваринам з внутрішнім ембріональним розвитком (ссавці) і тваринам з непрямим зовнішнім ембріональним розвитком (ланцетник).
Полілецитальні яйцеклітини містять багато жовтка. Вони властиві тваринам з прямим зовнішнім ембріональним розвитком (плазуни, птахи, яйцекладні ссавці). Яйцеклітини, що містять середню кількість жовтка, називають мезолецитальними. Такі яйцеклітини властиві тваринам із зовнішнім прямим і непрямим типом ембріонального розвитку (амфібії, більшість риб).
Жовток у цитоплазмі яйцеклітин розміщується неоднаково. В одних яйцеклітинах він розміщений рівномірно – ізолецитальні яйцеклітини (ссавці, ланцетник), в інших – локалізований у ділянці вегетативного полюса – телолецитальні яйцеклітини (амфібії, риби, плазуни, птахи та яйцекладні ссавці). Периферійний шар цитоплазми називають кортикальним. У цьому шарі жовтка немає і міститься значна кількість мітохондрій та кортикальних гранул. Останні є комплексами протеогліканів і глікопротеїдів. Через кортикальний шар відбувається перенесення поживних речовин, він бере участь у заплідненні та розвитку на найбільш ранніх етапах ембріогенезу.
Яйцеклітини вкриті трьома оболонками: первинною, вторинною й третинною. Первинна оболонка (жовткова, або вітелінова) – це власне плазмолема. Вторинна оболонка утворена фолікулярними клітинами та їхніми похідними. Останні, як правило, щільно прилягають до первинної оболонки і формують блискучу (прозору) зону. Фолікулярні клітини поблизу блискучої зони розміщені радіально. Вони та їх відростки, що пронизують блискучу зону, і контактують з первинною оболонкою утворюють так званий променевий вінець. Вторинна оболонка виконує трофічну та захисну функції, вона також запобігає поліспермії – проникненню багатьох сперматозоїдів у яйцеклітину. Найкраще розвинена ця оболонка у ссавців. Третинна обо-
53
лонка є лише в яйцеклітинах плазунів, птахів та яйцекладних ссавців. Її складові частини утворені секретом клітин яйцепроводів.
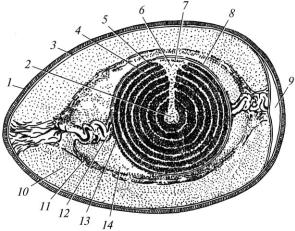
Будова яйцеклітини птахів. Яйцеклітину птахів називають яйцем (рис. 14). Яйце складається з жовтка, білка, підшкаралупової оболонки й шкаралупи. Жовток – це власне яйцеклітина, в якій є ядро і цитоплазма. Ядро знаходиться в ділянці анімального полюса і оточене вузькою смужкою цитоплазми, яка не має жовтка і містить органели (зародковий диск, рубчик). Жовток яйця неоднорідний, він буває світлим і темним. Поблизу центра яйцеклітини знаходиться світлий жовток. Ця зона має колбоподібну форму і називається латеброю. В інших ділянках яйцеклітини темний і світлий жовтки розміщені шарами. Темний жовток формується вдень, світлий – уночі. Жовток утримується в центрі за допомогою халаз – покручених пучків тонких ниткоподібних структур білково-вуглеводної природи, які спрямовані від жовтка до підшкаралупової оболонки протилежних кінців яйця. При цьому ядро яйцеклітини завжди знаходиться зверху. Вторинної оболонки у сформованій яйцеклітині птахів немає.
Рис. 14. Будова яйця курки (схема):
1 — шкаралупа; 2 — латебра; 3 — підшкаралупова оболонка; 4 — світлий жовток; 5 — темний жовток; 6 — безжовткова цитоплазма; 7 — ядро; 8 — жовткова оболонка; 9 — повітряна камера; 10 — білок (зовнішній шар); 11 — білок (волокнистий шар); 12 — халаза; 13 — халазоподібний шар білка; 14 — внутрішній шар білка
Білок, підшкаралупова оболонка і шкаралупа – це складові частини третинної оболонки. Часто окремо їх називають третинними оболонками. Білок розміщений навколо жовтка. До його складу входить вода (87%), білки, вуглеводи, мінеральні речовини та незначна кількість ліпідів. Він є поживним матеріалом для зародка. В білку є
54
також бактерицидні речовини. Залежно від локалізації розрізняють внутрішній і зовнішній шари білка. В кожному шарі білок буває в рідкому й щільному станах. Щільний білок бере участь у формуванні халаз.
Зовні білок вкритий підшкаралуповою оболонкою, яка складається з двох шарів – зовнішнього та внутрішнього. Внутрішній шар прилягає до білка, а зовнішній – до шкаралупи. В ділянці тупого кінця яйця між шарами оболонки знаходиться порожнина, заповнена повітрям – повітряна камера. Підшкаралупова оболонка побудована з волокон, до складу яких входять кератиноподібні речовини. Вона дуже щільна, еластична і проникна для газів, води та розчинів.
Шкаралупа побудована з неорганічних та органічних речовин. Органічні речовини утворюють щільно розміщені пучки волокон, а неорганічні – вапно. В шкаралупі є мікроскопічні пори, у зв’язку з чим вона проникна для газів. Зовні шкаралупа вкрита кутикулою, яка побудована із глікозаміногліканів. Вона запобігає проникненню крізь пори шкаралупи мікроорганізмів, спор грибів і проникна для газів. Шкаралупа виконує захисну функцію і є депо мінеральних речовин, які використовуються для формування скелета зародка.
Запитання для самоконтролю
1. Що вивчає ембріологія і як її ділять? 2. Чим відрізняються статеві клітини від соматичних? 3. Складові частини сперматозоїда. 4. Що знаходиться у головці сперматозоїда? 5. Що міститься в акросомі? 6. Що знаходиться в шийці сперматозоїда? 7. Які частини має хвіст сперматозоїда? 8. Що знаходиться у частинах хвоста сперматозоїда? 9. Чим відрізняються сперматозоїди ссавців і птахів? 10. Які функції виконує сперматозоїд? 11. Які особливості має яйцеклітина порівняно із сперматозоїдом? 12. Що таке полярна диференціація яйцеклітини? 13. Чим утворений жовток? 14. Які особливості кортикального шару цитоплазми яйцеклітини? 15. Скільки оболонок може мати яйцеклітина? 16. Чим утворена вторинна оболонка яйцеклітини ссавців, які функції вона виконує? 17. Чим утворений променевий вінець? 18. Критерії класифікації яйцеклітин. 19. Особливості будови яйцеклітини птахів. 20. Яка органела відсутня у яйцеклітині?
55
Лекція 7. Розвиток статевих клітин. Загальна характеристика ембріогенезу
Гаметогенез. Сперматогенез і овогенез. Запліднення. Зигота. Дроблення і його види. Бластула та її види. Гаструляція. Гаструла.
Онтогенез статевих клітин (гаметогенез). У ссавців первинні недиференційовані статеві клітини – гоноцити утворюються в стінці жовткового мішка зародка на ранніх етапах його розвитку. З течією крові або по ходу кровоносних судин вони мігрують у закладки статевих залоз (гонади), які називають статевими валиками. В останніх гоноцити стають нерухомими, округлюються, розмножуються і диференціюються в статеві клітини самця чи самиці. Диференціація відбувається залежно від типу статевих хромосом, якi були в клiтинах зародка. З диференцiацiєю статевих клітин починається і диференціація закладок статевих залоз.
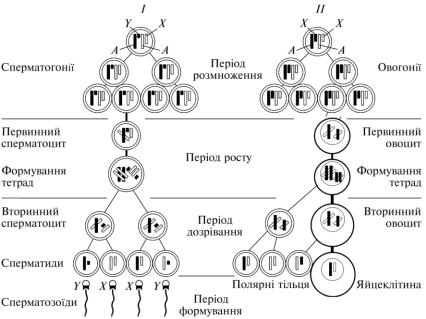
Сперматогенез – розвиток статевих клітин самця. Розвиток сперматозоїдів відбувається в статевих залозах самця – яєчках (сім’яниках) статевозрілих тварин. Він має чотири періоди: розмноження, росту, дозрівання і формування (рис. 15).
Рис. 15. Схема сперматогенезу (І) і овогенезу (ІІ):
А — аутосоми; Х, Y — статеві хромосоми
Період розмноження. У цей період статеві клітини називають сперматогоніями. Вони входять до складу стінки звивистих сім’яних канальців яєчок, мають невеликі розміри, плоску або округлу форму і
56
округлі ядра з диплоїдним числом хромосом. Сперматогонії активно діляться шляхом мітозу. Частина сперматогоній припиняє поділ, вони збільшуються в об’ємі і переходять у другий період розвитку. Клітини, які не припиняють ділитися, залишаються стовбуровими клітинами і поповнюють запас сперматогоній.
Період росту. Статеві клітини цього періоду називають первинними сперматоцитами. Вони втрачають зв’язок з базальною мембраною стінки звивистих сім’яних канальців і вступають у контакт з підтримувальними клітинами (клітини Сертолі). Ці клітини мають великі розміри і багато відростків. Вони виконують опорну, трофічну, захисну, секреторну та інші функції, створюючи потрібне середовище для статевих клітин, що розвиваються. В цей період у первинних сперматоцитах активно відбуваються процеси асиміляції, утворюються нові органели, що спричинює збільшення об’єму клітин. В ядрі синтезується ДНК і відбувається рекомбінація спадкового матеріалу (початок профази І мейозу). В ній виділяють п’ять стадій: лептотену, зиготену, пахітену, диплотену і діакінез. У лептотені відбувається конденсація хромосом. Вони стають помітними у ядрі і мають вигляд ниток. Хромосоми утворені двома хроматидами, які щільно прилягають одна до одної. Зиготена характеризується подальшою конденсацією хромосом, зближенням і з’єднанням (кон’югація) гомологічних хромосом. Кон’юговані хромосоми починають обмінюватись генами – кросинговер. У пахітені кон’юговані хромосоми потовщуються, стають короткими і тісно контактують по всій довжині. Для диплотени властиве формування тетрад. При цьому, у кожній кон’югованій хромосомі виникає щілина, яка розділяє хроматиди. Кон’юговані хромосоми починають відходити одна від одної, зберігаючи зв’язок у місцях перехресть. У діакінезі хромосоми потовщуються і відходять одна від одної.
Період дозрівання. В цей період відбуваються два мітотичні поділи клітин, які проходять один за одним з дуже короткою інтерфазою, у якій не відбувається подвоєння ДНК (мейоз). У результаті першого (редукційного) поділу з первинного сперматоцита утворюються два вторинних сперматоцити. Вони мають гаплоїдне число хромосом порівняно з первинними сперматоцитами, але кожна хромосома перед поділом складалась з двох хроматид, у зв’язку з чим їх набір залишається диплоїдним. Внаслідок другого (екваційного) поділу із вторинного сперматоцита утворюються дві сперматиди – істинно гаплоїдні клітини, які різняться за статевими хромосомами. Половина з них мають Х-, половина – Y-хромосому. Сперматиди – округлі клітини, перебувають у тісному контакті з підтримувальними клітинами.
57
Період формування. Упродовж цього періоду сперматиди набувають морфологічних ознак, властивих сперматозоїдам. При цьому ядро витягується, ущільнюється і переміщується до плазмолеми. Спереду від ядра із елементів комплексу Гольджі формується акросома. Клітинний центр переміщується до протилежного від ядра полюса клітини. В ньому розрізняють проксимальну та дистальну центріолі. Дистальна центріоль ділиться на краніальну й каудальну частини. Від краніальної частини формується аксонема, яка виходить за межі клітини. Каудальна частина набуває кільцеподібної форми. Вона «сповзає» по аксонемі разом з цитоплазмою сперматиди, яка містить мітохондрії та глікоген, розміщуючись на межі проміжної та головної частин хвоста. Сперматиди видовжуються і перетворюються на сперматозоїди. Частини цитоплазми сперматид, які не ввійшли до складу цитоплазми сперматозоїдів, фагоцитуються підтримувальними клітинами. Сперматозоїди втрачають зв’язок з підтримувальними клітинами і заповнюють просвіти звивистих сім’яних канальців.
Таким чином, при сперматогенезі з однієї сперматогонії утворюються чотири сперматозоїди.
Повний цикл розвитку сперматозоїдів кнура триває впродовж 40 діб, барана – 48, бугая – 63 діб.
Овогенез – розвиток статевих клітин самиць. Розвиток стате-
вих клітин самиць починається в ембріональний період і закінчується з настанням статевої зрілості. Він складається з трьох періодів – розмноження, росту та дозрівання (рис. 15). Перші два періоди відбуваються в статевих залозах самиць – яєчниках. Третій період починається в яєчниках і закінчується в яйцепроводах.
Період розмноження триває під час внутрішньоутробного розвитку і закінчується в перші місяці життя після народження. Статеві клітини цього періоду називають овогоніями. Овогонії – невеликі клітини округлої форми з оксифільною цитоплазмою і великим ядром, у якому міститься диплоїдне число хромосом. Вони активно розмножуються шляхом мітозу. Наприкінці періоду розмноження їхня мітотична активність припиняється і вони переходять у другий період розвитку. В період розмноження багато овогоній гине шляхом апоптозу.
Період росту – найтриваліший період розвитку, який закінчується з настанням статевої зрілості. Статеві клітини цього періоду називають первинними овоцитами, які знаходяться у фолікулах яєчника.
На початку періоду росту в ядрах первинних овоцитів відбувається рекомбінація спадкового матеріалу (профаза І мейозу) і фор-
58
мується вторинна оболонка яйцеклітини. Рекомбінація спадкового матеріалу відбувається так як і у періоді росту сперматогенезу, але остання фаза називається діктіотена. У цю фазу відбувається деспіралізація хромосом і вони стають невидимі до закінчення періоду росту. Вторинна оболонка утворена лише одним шаром фолікулярних клітин яєчників, у результаті чого утворюються примордіальні фолікули.
Після цього починається процес синтезу й накопичення у первинних овоцитах жовтка. Цей процес має дві фази – превітелогенезу та вітелогенезу і триває довго (роками, місяцями).
У фазі превітелогенезу (малого росту) в цитоплазмі первинних овоцитів збільшується кількість білоксинтезуючих органел і мітохондрій, які локалізуються переважно в периферійній частині цитоплазми. При цьому в ядрі овоцитів відбувається деспіралізація хромосом.
Вітелогенез (великий ріст) – фаза інтенсивного накопичення жовтка в цитоплазмі первинних овоцитів. Матеріал для синтезу жовтка надходить в овоцит зі всього організму через фолікулярні клітини, які збільшуються в об’ємі і активно діляться шляхом мітозу. Внаслідок цього вторинна оболонка стає багатошаровою (зернистий шар) і такі фолікули називають первинними. В міру накопичення жовтка ядро зміщується до одного з полюсів овоцита і утворюється кортикальний шар цитоплазми. З формуванням останнього процес синтезу й накопичення жовтка припиняється, в ядрі відбувається конденсація хромосом, ріст овоцитів закінчується. При цьому ріст первинних фолікулів триває. Між їхніми фолікулярними клітинами утворюються окремі порожнини, заповнені фолікулярною рідиною. Такі фолікули називають вторинними. Згодом окремі порожнини цих фолікулів з’єднуються, а первинний овоцит зміщується до стінки фолікула. Його місцезнаходження називають яйценосним горбком. Такі фолікули стають третинними (граафові міхурці).
Період дозрівання починається в яєчниках і закінчується після овуляції в яйцепроводах. У період дозрівання первинні овоцити діляться шляхом мейозу. В результаті першого поділу утворюються
вторинний овоцит і перше полярне тільце, яке майже не має цито-
плазми і містить половину хромосом. При наявності сперматозоїдів у яйцепроводі, один із них (ссавці) проникає у вторинний овоцит і стимулює другий поділ. У результаті його з вторинного овоцита утворю-
ються яйцеклітина і друге полярне тільце. Перше полярне тільце та-
кож може ділитись, унаслідок чого формуються два інших полярних тільця. При дозріванні яйцеклітина втрачає центріолі.
59
Таким чином, у результаті овогенезу з однієї овогонії утворюються одна яйцеклітина і три полярних тільця.
Полярні тільця та незапліднена яйцеклітина гинуть.
Запліднення та його біологічне значення. Запліднення – це процес злиття статевих клітин самця й самиці, в результаті якого утворюється одноклітинний зародок – зигота.
Запліднення поділяють на зовнішнє і внутрішнє. Зовнішнє запліднення відбувається за межами організму (ракоподібні, риби, амфібії), внутрішнє – в статевих органах самиці (птахи, ссавці).
Внутрішнє запліднення здійснюється в ампульній частині яйцепроводу. Воно відбувається в два етапи: дистантний і контактний. Дистантний етап передує власне заплідненню і починається ще в сім’явиносних шляхах самця перед еякуляцією (сім’явипорскуванням). У цей етап спочатку відбувається перебудова глікокаліксу плазмолеми сперматозоїдів, що захищає їх від руйнування в статевих шляхах самиці. Після потрапляння сперматозоїдів у статеві шляхи самиці вони вступають у контакт із секретом залоз та епітелію статевих шляхів, унаслідок чого здійснюється їх активація – капацитація. При цьому плазмолема акросомної ділянки сперматозоїдів звільняється від глікокаліксу і білків сперми, вона стає більш проникною для іонів кальцію, в результаті чого їх рухливість різко зростає. Завдяки своїм фізіологічним особливостям (реотаксис, хемотаксис) сперматозоїди досягають ампульної частини яйцепроводу, де знаходиться вторинний овоцит, і взаємодіють з ним (контактний етап запліднення). Цей етап починається з акросомної реакції. При цьому мембрана акросоми і плазмолема головки сперматозоїда зливаються, з акросоми виділяються гіалуронідаза й трипсиноподібні ферменти, які порушують контакти фолікулярних клітин вторинної оболонки і локально руйнують її прозору зону, внаслідок чого у цьому місці вторинний овоцит оголюється (денудація). Це дає можливість сперматозоїду проникнути в нього (пенетрація). Цитоплазма овоцита в місці контакту утворює горбок запліднення. На початку пенетрації плазмолема головки сперматозоїда і первинна оболонка овоцита зливаються і в цитоплазму овоцита проникає вміст сперматозоїда, тоді як його плазмолема залишається на поверхні овоцита. Після проникнення сперматозоїда у вторинний овоцит починається кортикальна реакція. При цьому з кортикального шару цитоплазми овоцита виходять біополімери, які взаємодіють з глікокаліксом первинної оболонки овоцита і утворюють непроникну для інших сперматозоїдів оболонку запліднення. Отже, кортикальна реакція забезпечує моноспермність
60
запліднення – проникнення в овоцит лише одного сперматозоїда. Моноспермія властива ссавцям.
Узаплідненому вторинному овоциті завершується другий поділ мейозу (у поділі бере участь тільки ядро), в результаті чого утворюються зріла яйцеклітина і одне полярне тільце. Ядро сперматозоїда перетворюється на пронуклеус самця, а яйцеклітини – на пронуклеус самиці. Вони зближуються, зливаються (синкаріон) і утворюють ядро зиготи, в якому міститься диплоїдне число хромосом. Хромосоми при злитті пронуклеусів формують метафазну екваторіальну пластинку. Центріоль, що вноситься в яйцеклітину сперматозоїдом, подвоюється. Центрiолi розходяться до полюсів зиготи і починає формуватись мітотичний апарат поділу. Таким чином, зигота вступає в наступний етап ембріонального розвитку – дроблення (дробіння).
Запліднення має велике біологічне значення. Завдяки цьому процесу здійснюється передавання спадкового матеріалу від батьків новому організму. В результаті виникає багато нових комбінацій спадкового матеріалу, утворюється різноманітний генофонд, який є матеріалом для добору в господарській діяльності людини.
Дроблення – це черговий етап ембріогенезу, який закінчується утворенням багатоклітинного зародка – бластули.
Після утворення зигота ділиться шляхом мітозу на дві клітини – бластомери, які не розходяться і продовжують ділитися разом. Поділи новостворених клітин відбуваються швидко, один за одним, внаслідок чого вони не досягають розмірів материнських. Такий процес поділу називають дробленням. Ядра дочірніх клітин не відрізняються від материнських, оскільки кожному поділу передує подвоєння ДНК. Співвідношення між ядрами й цитоплазмою бластомерів змінюється після кожного поділу, і коли воно набуває характерного для соматичних клітин певного виду тварин значення, дроблення закінчується.
Упроцесі дроблення спочатку утворюється група клітин, які щільно прилягають одна до одної. Далі вони зміщуються на периферію і формують пухирець з порожниною – бластулу.
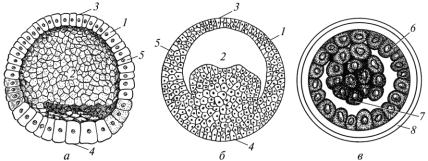
Бластула має кулясту форму і складається зі стінки (бластодерми) й порожнини (бластоцелі). Стінка утворена бластомерами. Вони щільно прилягають один до одного і розміщуються в один або кілька шарів. Бластодерма обмежує бластоцель – первинну порожнину тіла, заповнену рідиною. На бластулі розрізняють дах, дно і крайову зону
(рис. 16).
Залежно від кількості жовтка, який міститься в зиготі, швидкість
іхарактер дроблення неоднакові. У зв’язку з цим розрізняють два ви-
61
ди дроблення: повне (голобластичне) і часткове (меробластичне). Повне дроблення поділяють на рівномірне і нерівномірне. Повне рівномірне дроблення характерне для зигот з малим вмістом жовтка, який рівномірно розміщений у цитоплазмі (ланцетник). Під час цього дроблення в поділі бере участь уся зигота і бластомери мають майже однакові розміри. Для зигот із середнім вмістом жовтка, який сконцентрований у ділянці вегетативного полюса (окремі види риб, амфібії), характерне повне нерівномірне дроблення. У процесі цього
Рис. 16. Бластули хребетних тварин:
а — ланцетника; б — амфібії; в — ссавця; 1 — бластодерма; 2 — бластоцель; 3 — дах; 4 — дно; 5 — крайова зона; 6 — трофобласт; 7 — ембріобласт; 8 — прозора зона
дроблення в поділі бере участь уся зигота. Однак швидкість поділу вегетативного полюса, який переобтяжений жовтком, менша від швидкості поділу анімального полюса, внаслідок чого бластомери мають неоднакові розміри. У ділянці дна бластули вони великі (макромери), а в ділянці даху – малі (мікромери).
Часткове, або дискоїдальне дроблення характерне для зигот з великим вмістом жовтка (риби, плазуни, птахи). Дроблення в таких яйцеклітинах відбувається лише в ділянці анімального полюса. Частина зиготи, заповнена жовтком, у дробленні участі не бере.
Зиготи свійських ссавців містять малу кількість жовтка. Дроблення їх починається як повне рівномірне, далі воно відбувається як повне нерівномірне і асинхронне. Такий вид дроблення окремі автори визначають як неправильне або архаїчне (Токін Б.П., 1966).
Залежно від виду дроблення утворені бластули мають особливості будови та специфічні назви (рис. 16). У разі повного рівномірного дроблення утворюється одношарова, з великою порожниною бластула, яку називають целобластулою (ланцетник). У результаті повного нерівномірного дроблення (амфібії) формується багатошарова бластула – амфібластула (різновид целобластули). Її порожнина зміщена до даху. В разі часткового дроблення утворюється дискобластула
62