

Норм физ лекция двигательные системы (2 курс, 1 сем)
.pdf1. Общая организация двигательных функции ЦНС.
Периферическая часть двигательных систем - это нервно-мышечные синапсы и сами мышцы. Центральная часть представлена сегментарными и надсегментарными центрами. Аксоны нейронов сегментарных центром выходят из среднего, продолговатого и спинного мозга. Они непосредственно связаны с мышцами. Надсегментарные центры ( мозжечок, базальные ганглии, таламус) с периферией непосредственно не связаны. Аксоны их нейронов заканчиваются на клетках в других нервных центрах ( в том числе сегментарных).
Типы движений различаются по степени сложности и участия сознания в контроле выполняемых движений. В каждом из них есть 2 компонента: тонических ( обеспечивает необходимую для начала движения позу и на время фиксирует в начальном положении суставы) и фазный ( определяет направление и скорость движения).
Типы мышечной активности:
Локомоции - движения направленные на активное перемещение тела в пространстве. Формируются фазными сокращениями
Мышечный тонус - это постоянное непроизвольное напряжение мышц, необходимое для поддержания положения тела в пространстве. Поддерживается тонической активностью скелетных мышц.
Виды двигательной активности: 1) Элементарные движения 2) Активные движения (действия)
Функции двигательных систем регулируются по принципы обратной связи. Общий конечный путь всех двигательных систем - это альфа-мотонейроны, расположенные в передних рогах спинного мозга. Существуют 2 основных механизма управления движениями:
1) Сенсорная коррекция текущего движения по принципу обратной связи. 2) Прямое программное управление.
Двигательные программы запускаются много нейронными управляющими центрами, в которых обязательно есть клетки обладающие наименьшим порогом возбуждения. Эти клетки называются командные нейроны.
Схема управления движения
Двигательные системы построены по иерархическому принципу. Они условно делятся на 4 уровня. 1 - Двигательные системы спинного мозга, формирующие рефлекторные акты и элементарные движения.
2 - Двигательные системы ствола мозга ( продолговатый и средний мозг), которые обеспечивают цельные движения.
3 - Двигательные центры, осуществляющие коррекцию двигательных актов ( мозжечок и базальные ганглии)
4 - Это корковые структуры, которые отвечают за замысел движения и контролируют его.
2. Двигательные функции спинного мозга.
Основа двигательных систем спинного мозга - это соматические рефлекторные дуги, с участием которых реализуются двигательные рефлексы. Они подразделяются на 3 группы:
1.Собственные ( миотатические) рефлексы - рефлексы регуляции длины мышц - рефлексы регуляции напряжения мышц
2.Фазные рефлексы
-сгибательные
-разгибательные
-перекрестные сгибательные 3.Ритмические рефлексы
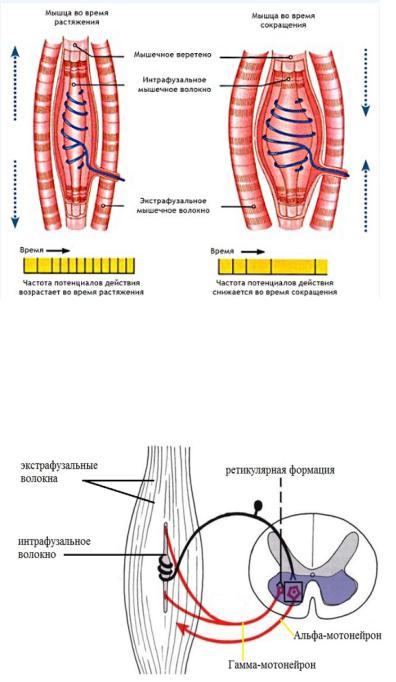
Рефлексы называются собственными, потому что отвечает та мышца, рецепторы которой раздражаются. Рефлекс регуляции длины мышц осуществляется с интрафузальных мышечных волокон. Это маленькие и тонкие волокна, которые находятся в толще поперечно-полосатых мышц. Они не участвуют в сократительном ответе, а выполняют исключительно рецепторную функцию. В середине каждого находится спирально закрученный рецептор. Он реагирует на растяжение всей мышцы или на растяжение самого интрафузального волокна.
Если вся мышца растягивается, то импульсы от спирально закрученного рецептора возбуждают альфа-мотонейрон, который иннервирует обычные поперечно-полосатые экстрафузальные волокна. В результате мышца сокращается..
Таким образом, мышца растягивается. возбуждается спирально-закрученный рецептор интрафузального волокна. Этот возбуждающий сигнал непрерывно доходит до альфа-мотонейрона и он вызывает сокращение всей мышцы.
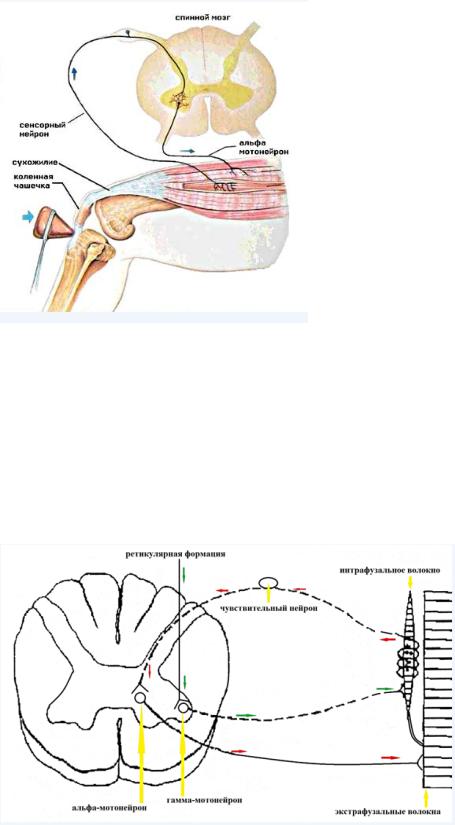
Коленный рефлекс
При ударе молоточком сухожилие деформируется, а мышца растягивается. Активирующий сигнал доходит до альфа-мотонейрона и вся мышца сокращается.
Все сухожильные рефлексы - это рефлексы с интрафузальных мышечных волокон.
Рефлекс с интрафузальных мышечных волокон, начинающийся с активации ретикулярной формации
Рефлекс с интрафузальных мышечных волокон может осуществляться и без предварительного растяжения всей мышцы. В этом случае он начинается с возбуждения ретикулярной формации, которая посылает активирующие сигналы на гамма-мотонейроны по ретикуло-спинального пути. Эти нейроны тоже находятся в передних рогах спинного мозга. Они иннервируют сократительных элементы концов интрафузального волокна. Когда концы интрафузального волокна сокращаются, его середина растягивается. При этом возбуждается спирально закрученный рецептор. Импульс доходит до альфа-мотонейрона и вся мышца сокращается, хотя исходно она находилась в состоянии покоя. Этот гамма-альфа рефлекс, начинающийся с активации ретикулярной формации и заканчивающийся сокращением скелетных мышц, является одних из компонентов первого вдоха новорожденного.
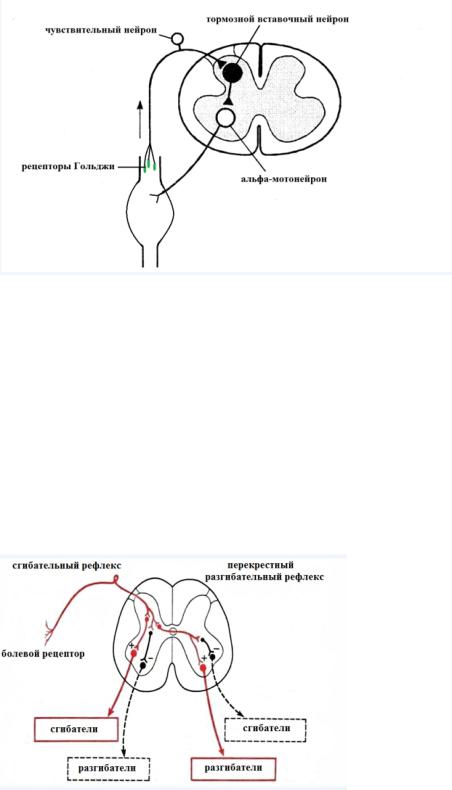
Рефлекс регуляции напряжения мышцы
Рефлекс осуществляется с рецептором Гольджи, которые находятся в сухожилиях. Началом рефлекса является сокращение мышцы. Когда мышца сокращается ее брюшко укорачивается, а сухожилия натягиваются. Натянутые сухожильные нити сдавливают рецепторы Гольджи и активируют их. Сигнал поступает в спинной мозг на тормозной вставочный нейрон, который блокирует активность альфа-мотонейрона, в результате чего мышца расслабляется.
Рефлекс рецепторов Гольджи выполняет защитную функцию в отношении двигательной системы. При длительном перенапряжение мышц активность альфа-мотонейронов тормозиться и мышцы непроизвольно расслабляются.
Фазные рефлексы спинного мозга Фазные рефлексы спинного мозга осуществляются с кожных рецепторов. К этим рефлексам относятся
сгибательные, разгибательные и перекрестные рефлексы, которые наблюдаются на противоположной конечности.
Сгибательные рефлексы возникают под влиянием потока импульсов, идущих от температурных или болевых рецепторов кожи. Эти сигналы возбуждают альфа-мотонейроны сгибателей и одноименной конечности. Одновременно они тормозят альфа-мотонейроны разгибателей это конечности, по типу реципрокного торможения. Сгибательные сложнее миотатических, тк одновременно в силу дивергенции импульсация от афферентных нейронов распространяется и на альфа-мотонейроны контралатеральной конечности. На противоположной стороне активируются альфа-мотонейроны разгибателей и одновременно тормозятся альфа-мотонейроны сгибателей. Следствием этого является то, что контралатеральная конечность выпрямляется, центр тяжести смещается в эту сторону, те рефлекс помимо уклонения от раздражителя способствует сохранению положения тела в пространстве.
Разгибательные и перекрестный рефлексы осуществляются с тактильных рецепторов.
Хорошо выражены у детей первых 3-х месяцев жизни и появляются у людей после травмы спинного мозга. У здоровых людей разгибательные рефлексы подавлены, потому что в целом преобладает тонус мышц-сгибателей.
Ритмические рефлексы спинного мозга ( шагатели и чесатели)
Шагательный рефлекс существует у детей первых 3-х месяцев жизни, а затем в норме подавляется супраспинальными механизмами. Он возникает при одновременном раздражении тактильных рецепторов обеих подошв. Нет классической рефлекторной дуги.
Чесательный рефлекс существует у всех животных и человека. Он представляет собой попеременное сгибание и разгибание конечности при раздражении тактильных рецепторов любого участка тела.
3. Двигательные функции ствола мозга
Врегуляции движений участвуют следующие структуры ствола мозга:
1)Двигательные ядра 3, 4 , 5, 7, 9, 10, 11 и 12 пар черепно-мозговых нервов ( жевание, глотание, артикуляция, движения глаза, смыкание век)
2)Вестибулярные ядра ( поддержание положения тела в пространстве, изменение тонуса мышц-антагонистов верхних и нижних конечностей).
3)Красное ядро ( активация альфа-мотонейронов сгибателей и торможение альфа-мотонейронов разгибателей)
4)Ретикулярная формация продолговатого мозга ( повышение тонуса мышц-сгибателей)
5)Ретикулярная формация моста ( повышение тонуса мышцразгибателей)
Таким образом ствол мозга обеспечивает поддержание равновесия в условиях покоя. Это проявляется в наличие статических и статокинетических рефлексов.
●Статические рефлексы - направлены на изменение тонуса скелетных мышц при изменение
положения тела в пространстве, а также перераспределения тонуса мышц с целью восстановления нормальной позы.
-познотонические - осуществляются с вестибулярных рецепторов и проприорецепторов шеи; центры рефлексов находятся в продолговатом мозге ( бульбарные рефлексы)
-выпрямительные - осуществляются с вестибулярных рецепторов и проприорецепторов шеи; центры рефлексов находятся в среднем мозге.
●Статокинетические рефлексы на сохранение равновесия и ориентации в пространстве при изменении скорости движения.
-лифтные - возникают при вертикально направленном ускорении
-транспортные - при горизонтальном направлении ускорении
-вращательные -при угловом.
Познотонические рефлексы представляют собой перераспределение тонуса мышц конечностей при изменении положение головы. Они осуществляются с вестибулярных рецепторов и проприорецепторов мышц шеи. При наклоне головы вперед увеличивает тонус мышц сгибателей на передних конечностей и тонус мышц разгибателей на задних. При запрокидывание головы назад, на передних конечностях увеличивается тонус разгибателей, при наклоне головы в сторону у животных увеличивается тонус разгибателей одноименной стороны на передних конечностях; у человека при наклоне головы так же перераспределяется тонус мышц, но только на нижних конечностях.
Статокинетические рефлексы существуют для удержания равновесия и поддержания положения тела в пространстве при действия ускорения.
При действие вертикально направленного ускорения возникают так называется, лифтные рефлексы (например. если движущийся вверх лифт резко останавливается, то у человека повышается тонус мышц сгибателей на ногах).
При действие горизонтально направленного ускорения возникают так называемые транспортные рефлексы, хорошо видны в движущемся транспорте ( например, когда автобус резко трогается с места у человека увеличивается тонус мышц разгибателей на ногах) .
При действие углового ускорения возникает вестибуло-глазодвигательный и вестибуло-шейные рефлексы. Эти рефлексы представляют собой движение глаз и головы в стороны противоположную вращению, с последующим быстрым возвратом в исходное положение.
4. Двигательные функции мозжечка
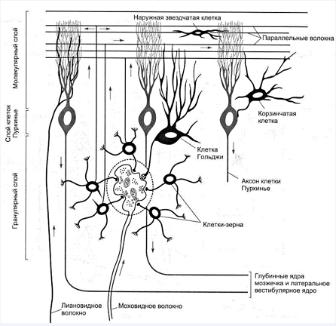
К коре мозжечка подходит 2 виды волокон: лиановидные и моховидные.
Лиановидные волокна являются аксонами нейронов нижней оливы продолговатого мозга. Информация к ним поступает от проприорецепторов, кожных рецепторов и двигательной коры головного мозга. Каждая лиановидная волокно устанавливает контакт с дендритами 1 клетки Пуркинье, но число синапсов может достигать 300, что объясняет исключительно сильное возбуждающее действие этого афферентного хода. По моховидным волокнам в кору мозжечка поступает информация от коры больших полушарии, от ретикулярной формации, проприорецепторов и вестибулярных рецепторов. Моховидные волокна образуют синапсы на дендритах клеток зерен внутреннего слоя коры. Активность клеток зерен регулируется через тормозные клетки Гольджи внутреннего слоя по типу возвратного торможения. Здесь используется медиатор - ГАМ-кислота.
Клетки Пуркинье. образующие средний слой являются главной функциональной единицей мозжечка с ярко выраженной интегративной направленностью. Эти клетки имеют сильно ветвящиеся дендриты, на которых находятся свыше 100 тыс синапсов. Клетки являются единственными эфферентными нейрона в коре мозжечка. Они связывают кору мозжечка с его ядрами и с вестибулярными ядрами среднего мозга. Клетки Пуркинье получают в основном возбуждающие сигналы, но сами посылают только тормозные сигналы к 3 парам мозжечковых ядер.
Мозжечок - это единственная структура головного мозга, в которой так сильно представлены процессы торможения. Каждый возбуждающий сигнал поступивший в мозжечок минимум через 2 синапса превращается в тормозной импульс, максимум через 4 синапса. такое разлитое торможение необходимо, для управления программами быстрых движений, поскольку информация о предыдущем этапе движения должна мгновенно стираться.
Регуляция мышечного тонуса обеспечивающего поддержания равновесия осуществляется преимущественно вестибулярной и спинальной частями мозжечка. Получая и обрабатывая импульсы от вестибулярных рецепторов ( кожных, зрительных) мозжечок оценивает состояние мышц, положение тела в пространстве, через ядра шатра и также посредством прямого выхода на вестибулярные ядра. Координация движении осуществляется в спинальной части мозжечка и его полушарий. В кору мозжечка поступают сигналы и от рецепторов мышц, связок и сухожилий и от моторной коры. Анализируя информацию о программе и ходе выполнения движения мозжечок через свои промежуточные ядра, имеющие выходы на красное ядра и моторную кору, осуществляет координацию, а
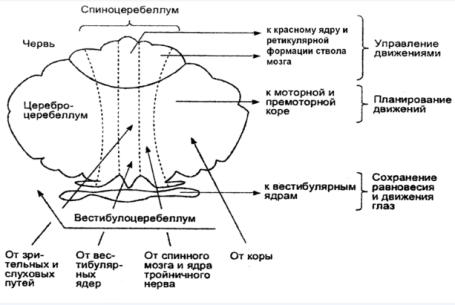
при необходимости и коррекцию движений. Программирование произвольных движений осуществляется только корой полушарий мозжечка, которая получает информацию о замысле движения их ассоциативных зон коры больших полушарий, через ядра моста. В коре мозжечка она перерабатывается в программу движения, которая через зубчатое ядро и ядра таламуса поступает в премоторную зону коры больших полушарии, где она получает дальнейшую обработку и реализуется как сложное целенаправленное движение. Коррекция и координация медленных движений осуществляется на основе обратной афферентации от поприорицепторов, а коррекция быстрых движений происходит сразу, путем прямого изменения программы.
Функции мозжечка:
1)Регуляция позы, мышечного тонуса и равновесия
2)Координация целенаправленного движения с рефлексами
3)Коррекция медленных точных движений
4)Управление содружественными движениями
5)Организация быстрых целенаправленных движений
6)Создание и хранение приобретенных двигательных программ ( ходьба, бег, игра на музыкальных инструментах, умение кататься на велосипеде)
7)Торможение миотатических спинальных рефлексов регуляции длины и напряжения мышц
Симптомы поражения мозжечка
Выделяют три основных симптома поражения мозжечка, которые в физиологии называются триада Лучиани(?). К ней относятся следующие симптомы:
1)Атония - отсутствие мышечного тонуса
2)Астазия - нарушение равновесия, качательные движения в вертикальном положении, тремор, усиливающийся в конце движения.
3)Астения - быстрая утомляемость мышц при выполнении любого движения.
Далее добавлены следующие:
4)Атаксия - нарушение равновесия, точности и координации движения.
++++
5)Асинергия - нарушение содружественных движений
6)Дизартрия - расстройство артикуляции
7)Дисметрия - утрата соразмерности движения, что особенно наблюдается при совершении целенаправленного движения.
8)Адиадохокинез - неспособность быстро и равномерно выполнять противоположные движения
* Выполнение пальценосовой пробы при повреждении мозжечка очень затруднено, поскольку мозжечок контролирует точность движения. Важно отметить, что с течением времени мозжечковые синдромы
сглаживаются, поскольку функции управления координации движения частично берут на себя другие отделы двигательной системы.
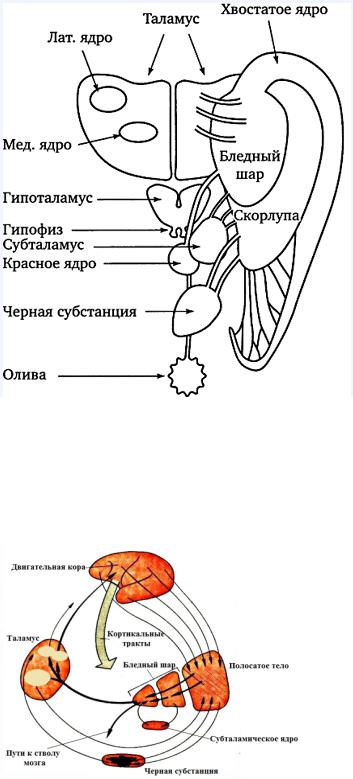
5. Двигательные функции базальных ганглиев
К базальным ганглиям относятся:
● Полосатое тело, состоящее из хвостатого ядра и скорлупы
● Бледный шар ● Субталамическое ядро
Они получают сигналы из двигательной области коры головного мозга. Базальные ганглии напрямую связаны со структурами среднего мозга ( двигательными ядрами таламуса, красным ядром, черной субстанции и ретикулярной формацией). Через связи с этими структурами базальные ганглии могут влиять на кортико-спинальный, рубро-спинальный и вентромедиальный нервные пути. Базальные ганглии играют главную роль в процессе перехода от замысла движения к выбранной программе действия.
Сигнал о замысле движения получает полосатое тело. Далее оно обменивается информацией с черной субстанцией. нейроны
черной субстанции синтезирует медиатор дофамин, который притормаживает активность полосатого тела. После этого полосатое тело отправляет возбуждающие сигналы в бледный шар. Бледный шар обменивается этой инофрмацией с субталамическим ядром, а затем отправляет его в моторные ядра таламуса. Талмус возвращает переработанную в базальных ганглиях программу движений обратно в двигательную кору. Информация о конкретных движениях может циркулировать в структурах базальных ганглиях очень долго ( до нескольких часов).
В этих взаимосвязях базальных ганглиях различают 2 петли циркуляции информации ( большую и маленькую) .
Большая начинается в двигательной коре →далее информация поступает в полосатое тело →затем сигналы в черную субстанцию, получая обратно тормозные в виде медиатора ( дофамина) → полосатое тело активирует бледный шар ( нейроны полосатого тела выделяют медиатор - ацетилхолин) →бледный шар обменивается информацией с субталамическим ядром, затем направляет сигналы в таламус → таламус возвращает программу движений в двигательную кору.
Для циркуляции импульса, для организации медленных и плавных движений существует малая петля.
Она начинается в полосатом теле → полосатое тело обменивается информацией с черной субстанцией, отправляя сигналы в бледный шар → бледный шар тормозит таламус → таламус посылает сигналы снова в полосатое тело. Импульсы могут циркулировать несколько часов.
Основные функции базальных ганглиев:
1)Приведение двигательной системы в готовность перед началом движения
2)Инициация двигательного акта, те организация первых этапов выполнения движения
3)Придание плавности быстрым движениям
4)Организация медленных движений
5)Облегчение запускаемых корой движений и подавление лишних сопутствующих движений
6)Базальные ганглии являются хранилищем врожденных стереотипных двигательных программ.
Симптомы поражения базальных ганглиев связаны с тем - функция каких именно частей системы нарушены. В целом, при повреждении какого-либо отдела базальные ганглии начинают навязывать свои тип движения всей двигательной системы.
●Поражение полосатого тела: - Гипертонус скелетных мышц
- торможение формирование условных рефлексов - появление навязчивых купноразмашистых движений
●Поражение бледного шара
-пластическая ригидность мышц
-медленные червеобразные движения пальцев ● Разрушение нейронов черной субстанции:
-затруднение начала движения
-гипокинезия( амимия, шаркающая походка)
-тремор покоя
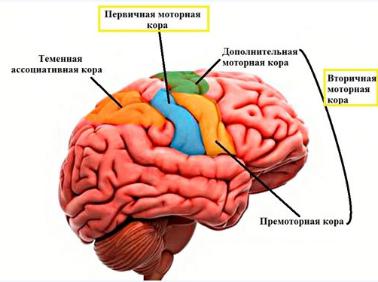
6. Роль коры больших полушарий в регуляции движения
В коре находятся первичная и вторичная моторная кора.
Первичная моторная кора располагается в передней центральной извилине. Афферентные сигналы она получает от соматосенсорной коры, премоторной коры, таламуса и мозжечка. Эфферентные сигналы кора отправляет к нейронам ядер черепно-мозговых нервов, красного ядра и ретикулярной формации.
Первичная моторная кора содержит гигантские пирамидные клетки Бетса (?). Их аксоны и формируют пирамидные кортико-бульбарные тракты. Пирамидный путь - это единственная прямая связь
головного и спинного мозга. В пирамидном тракте находится около 1 млн. нервных волокон.
Вторичная моторная кора включает в себя области премоторной и дополнительной моторной коры. Премоторная кора играет основную роль в программировании и организации движении, находящихся под контролем зрения, кроме того она участвует в организации позы. повреждение премоторной коры часто вызывает тенденцию повторного выполнения начатого движения ( персеверация ). Дополнительная моторная кора отправляет сигналы в первичную моторную кору, в спинной мозг и стволовые двигательные ядра. Кора принимает участие в формирование программы предстоящих сложных движении.
Симптомы поражения первичных моторной коры и пирамидного тракта.
1)Гемиплегия - парез или паралич мышц одной половины тела)
2)Исчезновение или резкое ограничение произвольной двигательной активности.
3)Гипертонус мышц
4)Усиление миотатических рефлексов, исчезновение фазных рефлексов
5)Отсутствие брюшных рефлексов
6)Появление мышечного сопротивления пассивным движениям по типу складного ножа.
7)Повышение тонуса сгибателей рук и разгибателей ног