

Генетический код и-рнк
первый нуклеотид триплета (5 – конец) |
второй нуклеотид триплета |
третий нуклеотид триплета (3-конец) |
|||
|
а |
г |
у |
ц |
|
а |
лиз лиз асн асн |
арг арг сер сер |
иле мет иле иле |
тре тре тре тре |
а г у ц |
г |
глу глу асп асп |
гли гли гли гли |
вал вал вал вал |
ала ала ала ала |
а г у ц |
у |
стоп стоп тир тир |
стоп три цис цис |
лей лей фен фен |
сер сер сер сер |
а г у ц |
ц |
глн глн гис гис
|
арг арг арг арг |
лей лей лей лей |
про про про про |
а г у ц |
СИНТЕЗ БЕЛКА включает несколько уровней: 1. Претранскрипционный
2. Транскрипционный 3. Транспортный 4. Трансляционный 5. Посттрансляционный. (рис. 13).
1. Претранскрипционный уровень: стартовый этап синтеза. Во время него происходит активация молекулы ДНК с помощью специальных белков.
2. Транскрипционный уровень: Транскрипция - синтез и-РНК на молекуле ДНК с последовательностью нуклеотидов комплементарной молекуле ДНК. Матрицей для и-РНК служит только одна из двух имеющихся цепей ДНК, то есть синтез носит полуконсервативный характер. Процесс контролируется ферментами РНК-полимеразами.
В молекуле ДНК выделяется определенный фрагмент- промотор как начальный участок синтеза. За ним следуют информационные нуклеотидные кодоны. Существует 2 вида наборов нуклеотидов. Триплеты, несущие информацию об аминокислотах – экзоны. Они образуют короткие сегменты ДНК и, как правило, разделены более длинными неинформативными участками нуклеотидов, которые называют интронами. В каждом гене количество экзонов на один превышает число интронов. С началом транскрипции в первичную и-РНК включаются как экзоны, так и интроны. Первичный транскрипт полностью копирует молекулу ДНК.
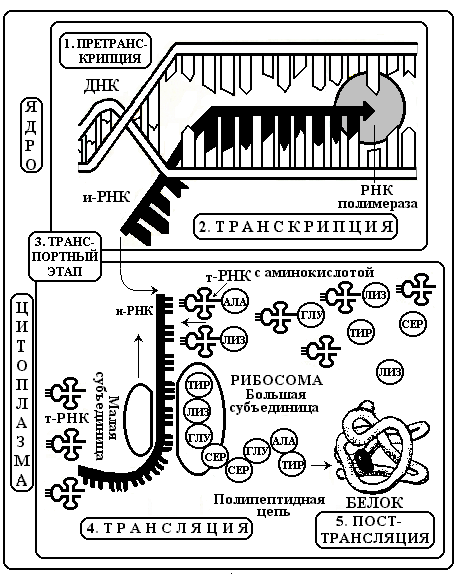 Рис.
13. Схема этапов синтеза белка
Рис.
13. Схема этапов синтеза белка
3. Транспортный уровень: охватывает период между транскрипцией и трансляцией. На данном этапе происходит процессинг, то есть созревание и-РНК. Суть его – удаление интронов. Экзоны сохраняются и соединяются в единую цепь с помощью ферментов лигаз. Указанное явление называется сплайсинг. В ходе его возможны ошибки транскрипции, когда удаляются не все интроны. Прошедшая сплайсинг и-РНК значительно укорочена, компактна и может служить матрицей для синтеза белка. Для этого она переносится из ядра в цитоплазму к рибосомам. В клетках животных и растений на всем пути от ядерного хроматина к рибосомам и-РНК находится не в свободном состоянии, а связана со специальными белками-переносчиками, часть из которых различна в ядре и цитоплазме. Белки «везут и-РНК в рибосому на перекладных».
4. Трансляционный уровень: Трансляция – синтез полипептидной цепи из аминокислот согласно кодирующей и-РНК. В ходе трансляции происходит перевод генетической информации в аминокислотную последовательность: ДНК, и-РНК, белок. Трансляция является очень важной частью общего метаболизма клетки. В ней задействованы не менее 20 ферментов (аминоацилсинтетаз), до 60 различных т-РНК, 3-5 молекул р-РНК и макромолекулы и-РНК. Здесь выделяют следующие этапы: инициация, элонгация, терминация.
Инициация – начало трансляции. Фермент аминоацилсинтетаза активирует определенную аминокислоту и вовлекает ее в белковый синтез. Активированная аминокислота соединяется с т-РНК специализированной на данной аминокислоте. Доставленная к рибосоме и-РНК связывается с рибосомальной РНК. При этом рибосома диссоциирует, ее малая и большая субъединицы несколько раздвигаются, и рибосома насаживается на и-РНК. В начальной части и-РНК имеются стартовые кодоны АУГ и ГУГ. К ним присоединяется т-РНК, несущая соответствующую аминокислоту формилметионин. Следовательно, эта аминокислота всегда является первой в синтезируемом на рибосоме белке.
Элонгация – продолжение трансляции. Рибосома двигается вдоль молекулы и-РНК. Транспортные РНК связываются с рибосомой и их антикодоны избирательно по правилу комплементарности контактируют с кодонами и-РНК. При совмещении кодона и антикодона аминокислота отрывается от т-РНК и включается в полипептидную цепь из аминокислот в большой субъединице рибосомы. При формировании первичной белковой структуры каждая новая аминокислота занимает место за аминокислотой, включенной непосредственно перед ней.
Терминация –окончание синтеза. В и-РНК есть трейлерный участок, содержащий стоп-кодоны УАА, УАГ, УГА. Они указывают на завершение синтеза данного белка. Поэтому при контакте с ними сборка аминокислотной цепи заканчивается
5. Посттрансляционный.уровень: синтезированная белковая молекула начинает усложнять свою первичную структуру под действием ферментов. Происходит ее конформация, изменяется пространственная организация белка, он приобретает вторичную, третичную и четвертичную структуры.
Таким образом, мы познакомились с основными этапами синтеза белковой молекулы и узнали центральную догму молекулярной биологии. Она гласит: информация в живых организмах передается по цепи ДНК-РНК-белок. В настоящее время доказано явление обратной транскрипции, когда передача информации происходит от РНК к ДНК. В то же время совершенно невозможен перенос информации от белков обратно к нуклеиновым кислотам. Это означает, что изменения (модификации) белков, как конечных генных продуктов, не наследуются.
В соответствии с существующими представлениями синтез белка всегда начинается с работы генов. Под ней подразумевается их способность транскрибировать, то есть направлять синтез и-РНК. Но не всегда работа гена заканчивается сборкой белковой молекулы. Как заметил один американский генетику, для того, чтобы выдать белок гену нужно пробиться через «клеточные джунгли». Выход конечного белкового продукта – это экспрессия гена. Она совершается в результате деятельности всей клетки с ее многокомпонентными механизмами белкового синтеза.
ОПЕРОННАЯ СИСТЕМА:
Гены функционально неоднородны. Это обнаружили в 1961 году французские ученые Ф. Жакоб и Ж. Моно. Они доказали, что существует две группы генов: структурные, управляющие синтезом специфических белков и регуляторные, контролирующие деятельность структурных генов. Синтез белка в клетке регулируется с помощью оперонной системы, состоящей из регуляторных (промотор и оператор) и структурных генов (рис. 14).
Следовательно, регуляция белкового синтеза в организме эукариот — процесс очень сложный. Он связан с наличием в клетке и с существованием, помимо генной, нервно-эндокринной системы регуляции. Открытия, сделанные в области молекулярной генетики, являются величайшими достижениями биологии.
Рис.
14. Схема оперонной системы регуляции
синтеза специфического белка у прокариот.