

8. Нуклеопротеиды. Химический состав белковой и простетической группы. Структурные компоненты нуклеиновых кислот. Номенклатура нуклеотидов, нуклеозидов, азотистых оснований. Их химическое строение.
Нуклеопротеиды – это сложные белки, содержащие в качестве небелковой части нуклеиновые кислоты (до 65%).
НП состоят из 2-х частей: белковой (содержит гистоны и протамины, которые являясь основными белками, придают основные свойства) и простетической, представленной НК, сообщающими кислотные свойства. Взаимодействие между этими частями по ион-ионному механизму.
Все НП по составу НК можно разделить на 2 группы: рибонуклеопротеиды (РНП) и дезоксирибонуклеопротеиды (ДНП).
Состав нк:
НК – высокомолекулярные органические вещества, полинуклеотиды. Мономерами являются мононуклеотиды. Каждый мононуклеотид состоит из: углевода, азотистого основания и фосфорной кислоты. Так, РНК содержит -D-рибофуранозу (рибозу), одно из 4-х возможных азотистых оснований (А, Г, Ц или У) и остаток фосфорной кислоты. ДНК содержит -D-дезоксирибофуранозу (дезоксирибозу), одно из 4-х возможных азотистых оснований (А, Г, Ц или Т) и остаток фосфорной кислоты.
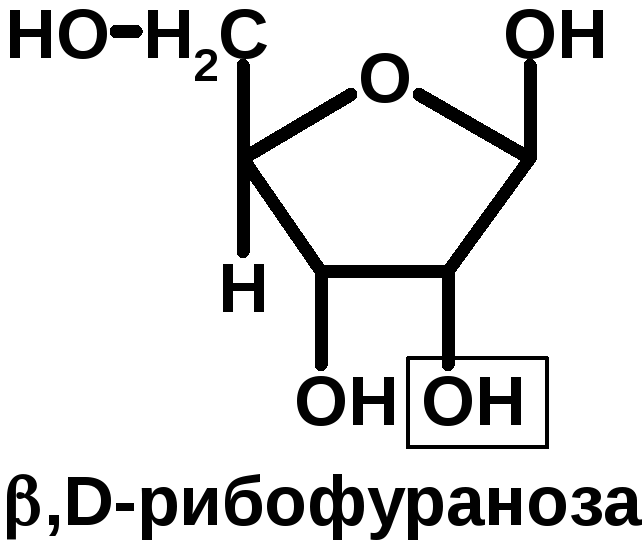
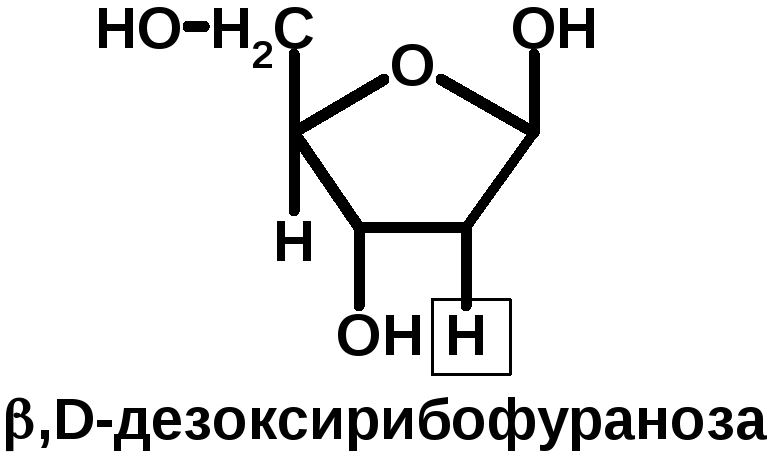
Строение азотистых оснований:
К группе пуриновых относятся аденин (6-аминопурин) и гуанин (2-амино-6-оксипурин). К группе пиримидиновых – урацил (2,4-диоксипиримидин), тимин (5-метилурацил) и цитозин (2-окси-4-аминопиримидин).
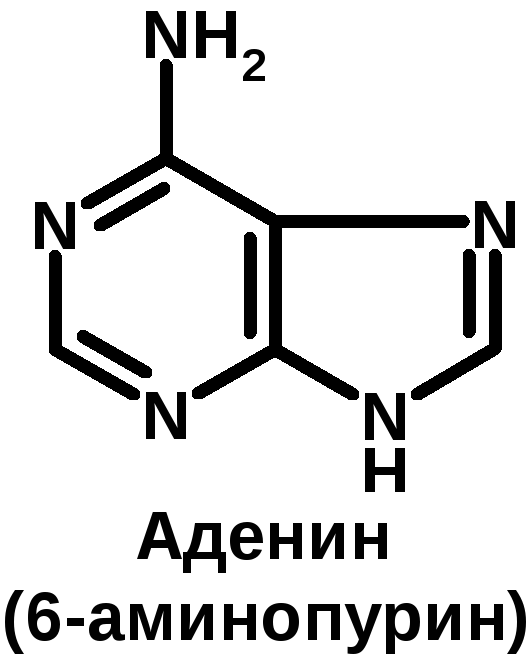
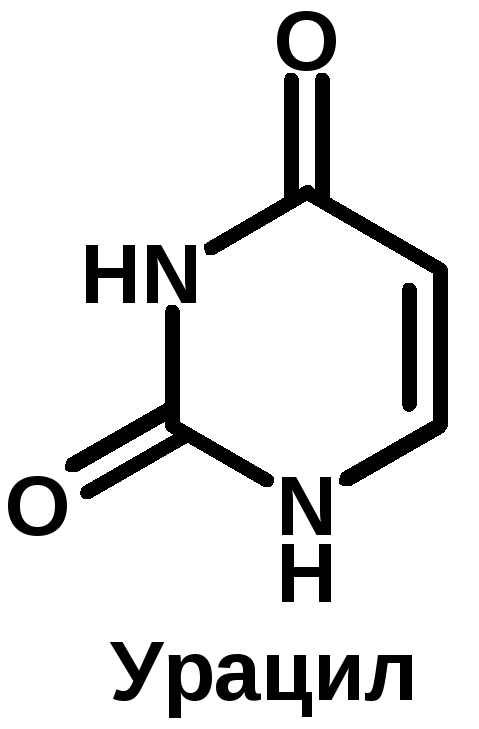
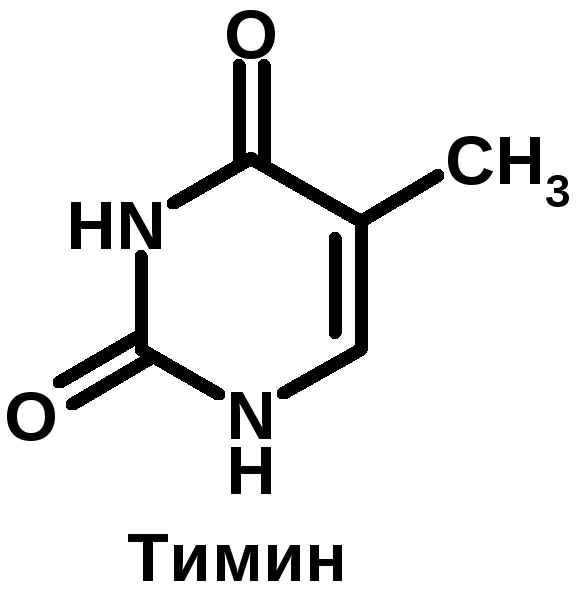
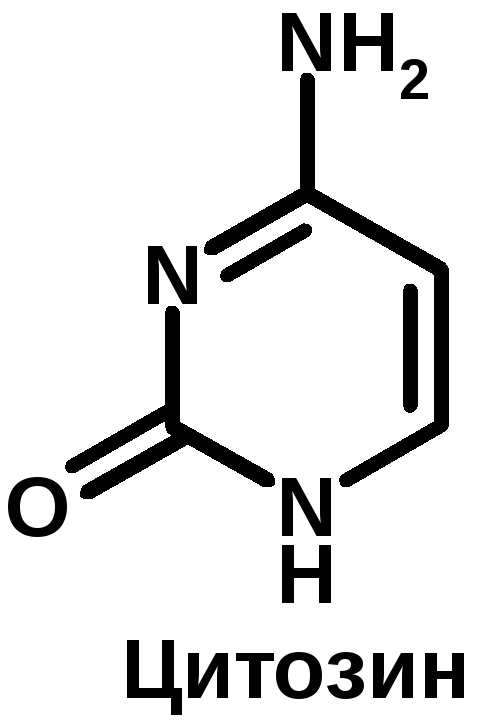
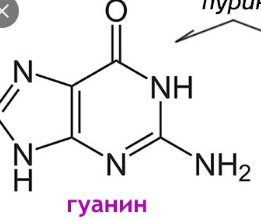
Структура нуклеиновых кислот:
Имеют несколько уровней структурной организации.
1. первичная структура. РНК и ДНК построены однотипно – представлены полинуклеотидной цепью, состоящей из отдельных мононуклеотодов, соединённых между собой 3’→5’-фосфодиэфирными связями. Эта связь образуется между фосфорным остатком одного мононуклеотида и 3’-ОН-группой пентозного остатка другого мононуклеотида. [рис. образования такой связи] Разные НК отличаются числом, порядком чередования и составом НК.
2. вторичная структура. По рентгеноструктурному анализу ДНК в 1953г Уотсон и Крик предложили модель строения ДНК, которая объясняла самовоспроизведение организмов, наследственную изменчивость. Вторичная структура представляет собой двойную спираль, состоящую из 2 полинуклеотидных цепей, закрученных вокруг одной общей оси. Эти цепи антипараллельны, т.е. одна идет в направлении 5’→3’, а другая 3’→5’. Пуриновому основанию одной цепи соответствует пиримидиновое основание другой цепи – эти основания комплиментарны друг другу, т.е. дополняют одно другое до целого. Между А и Т две водородные связи (А=Т), а между Г и Ц – 3 (ГЦ).
Молекула спирализована на всем протяжении, гидрофобные участки внутри спирали, их плоскости перпендикулярны основаниям и параллельны друг другу. В вертикальном направлении возникают гидрофобные взаимодействия. Вторичная структура стабилизируется водородными связями и гидрофобными взаимодействиями.
Вторичная структура РНК более простая, представляет собой одну полинуклеотидную цепь, в которой спирализованы лишь некоторые участки. Вторичная структура РНК представлена в виде клеверного листа. Для тРНК известна третичная структура в форме буквы Г. [рис. РНК в виде клеверного листа]
9. Гемоглобин. Строение и свойства. Окси-, карбокси-,карб- метгемоглобин. Вариации первичной структуры и свойства гемоглобина. Гемоглобинопатии.
Гемоглобин (Hb)- состоит из белка – глобина и железосодержащего гема. Глобин - белковый компонент, гем – небелковый. Гемоглобин относится к хромопротеидам. (Красный цвет придаёт железо). Видовые различия обусловлены глобином. Гем одинаков у всех видов Hb.
Гем- комплекс железа с порфирином. В основе структуры порфиринов лежит порфириновое кольцо, состоящее из 4-х пирольных колец. В природе порфирины входят в состав хлорофилла. Порфирины способны к флуоресценции.
Структура гемоглобина. Каждая молекула Hb состоит из 4-х субъединиц (протомеров), включающих гем и глобин. Протомеры соединены гидрофобными, ионными, водородными связями. Структура Hb обеспечивает наиболее удобное связывание кислорода.
Биологическая роль Hb . Hb находится в эритроцитах, переносит кислород от лёгких к тканям и углекислый газ от тканей к лёгким. Способен связывать углекислый газ, угарный газ, цианиды. Связывая протоны Н+, поддерживает постоянство РН в эритроцитах, выступая в роли внутриклеточного буфера. В 1 эритроците содержится 340 000 000 молекул Hb.

Гемоглобин, присоединяя некоторые молекулы или ионы образует несколько форм:
HbО2 (Fe2+) – окисленный (оксигемоглобин). Основная форма гемоглобина в артериальной крови -96-98%.
НHb (Fe2+) – восстановленный (дезоксигемоглобин). Содержится в венозной крови.
HbСО - карбоксигемоглобин. Образуется путём присоединения к Hb окиси углерода (СО угарного газа). Не способен переносить кислород . У здоровых – 0,25-2%, у новорожденных – 10-12%. Уровень СО Hb зависит от экологических факторов, вредных привычек, характера работы… У курильщиков – 7-10%, у таксистов – до 20%, при эпилепсии, коме, смерти – более 50%.
HbCO2 – карбогемоглобин, образуется при связывании углекислого газа в тканях, в венозной крови составляет 15-20% от всего количества гемоглобина.
HbОН – метгемоглобин (Met Hb), содержит Fe 3+. Не способен переносить кислород. Появляется в крови при отравлении оксидами азота, нитробензолом, анилиновыми красителями. Концентрация в крови более 70% - смерть. Помощь – вдыхание чистого кислорода.
Вариации первичной структуры:
Hb взрослого организма состоит, как упомянуто выше, из двух α- и двух β-цепей (α2β2). Наряду с этой основной формой (HbA1) в крови присутствуют незначительные количества второй формы с более высоким сродством к O2, у второй β-цепи заменены δ-цепя-ми (HbA2, α2δ2). Две другие формы Hb встречаются только в эмбриональном периоде развития. В первые три месяца образуются эмбриональные гемоглобины состава α2ε2-P и α2γ2-F. Затем вплоть до рождения доминирует фетальный гемоглобин (HbF), который постепенно заменяется на первом месяце жизни на HbА. Эмбриональный и фетальный гемоглобины обладают более высоким сродством к О2 по сравнению с HbА, так как они должны переносить кислород из системы материнского кровообращения.
Гемоглобинопатии (от гемоглобин и греч. páthos — страдание, болезнь), гемоглобинозы, состояния, обусловленные присутствием в красных кровяных тельцах (эритроцитах) одного или нескольких аномальных (патологических) гемоглобинов. Выделено свыше 50 патологических разновидностей гемоглобина, возникших в результате врождённого, передаваемого по наследству дефекта образования белковой части гемоглобина — глобина. При аномалиях гемоглобина нарушаются физико-химические свойства эритроцитов, обменные процессы в них; эритроциты становятся менее устойчивыми к различным гемолизирующим факторам. Наиболее распространены и отличаются тяжестью проявлений серповидноклеточная (дрепаноцитарная) анемия и талассемия. Серповидноклеточная анемия (HbS) связана с наличием в эритроцитах патологического гемоглобина S (первая буква англицская side — серп). При этой форме Г. эритроциты в условиях снижения парциального давления кислорода в окружающей среде приобретают форму серпа. При увеличении в крови количества серповидных эритроцитов нарастает вязкость крови, замедляется кровоток, происходит разрушение серповидных эритроцитов, развиваются тромбозы в различных органах. У практически здоровых носителей HbS серповидность эритроцитов и появление признаков заболевания могут наступить лишь в условиях гипоксии. Поэтому всем носителям HbS противопоказаны служба в авиации, а также полёты на самолётах без достаточного кислородного обеспечения. Талассемия — заболевание, распространённое в средиземноморских странах. Характеризуется значительным повышением содержания HbF в крови. Полагают, что при этом образование нормального гемоглобина HbA подавлено. Нарушено также образование железосодержащей части гемоглобина (гема). Различают большую, малую и минимальную талассемию. Для всех форм талассемии характерно наличие в крови "мишеневидных" эритроцитов, в которых гемоглобин расположен в центре клетки в виде мишени.