

64. Апотомический распад углеводов. Биологическое значение пентозофосфатного цикла.
 Пентозофосфатный
путь (ПФП)
Наиболее активно реакции ПФП идут в
цитозоле клеток печени, жировой ткани,
эритроцитах, коре надпочечников, молочной
железе при лактации, в гораздо меньшей
степени в скелетных мышцах. Этот путь
окисления глюкозы не связан с образованием
энергии, а обеспечивает анаболизм
клеток. (у детей первых лет жизни
активность ПФП высокая).
Пентозофосфатный
путь (ПФП)
Наиболее активно реакции ПФП идут в
цитозоле клеток печени, жировой ткани,
эритроцитах, коре надпочечников, молочной
железе при лактации, в гораздо меньшей
степени в скелетных мышцах. Этот путь
окисления глюкозы не связан с образованием
энергии, а обеспечивает анаболизм
клеток. (у детей первых лет жизни
активность ПФП высокая).
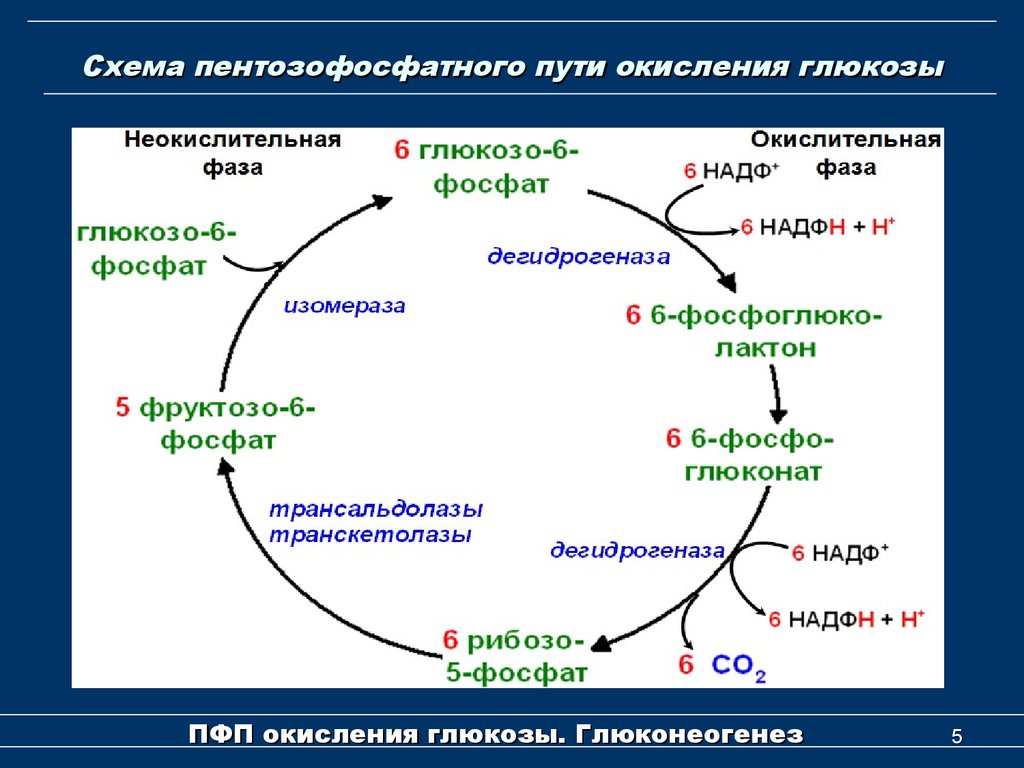
Пентозофосфатный путь включает два этапа – окислительный и этап структурных перестроек (неокислительный).
На первом этапе (окислительном) глюкозо-6- фосфат превращается в рибулозо-5-фосфат, реакции сопровождаются восстановлением двух молекул НАДФ до НАДФН. На этом этапе происходит регуляция процесса: инсулин повышает активность глюкозо-6- фосфатдегидрогеназы и фосфоглюконат-дегидрогеназы.
Первая реакция. Катализатор — глюкозо-6-фосфатдегидрогеназа. Кофермент реакции НАДФ. В ходе этой реакции от молекулы глюкозо-6-фосфата отщепляется два водорода. При этом НАДФ присоединяет к себе гидрид ион. Гидрид ион состоит из протона водорода и двух электронов. Образуется глюконолактон-6-фосфат и НАДФH. Один протон отправляется в цитоплазму.Вторая реакция. Фермент — глюконолактон-гидратаза. У неё есть название попроще — лактоназа. Лактоназа присоединяет молекулу воды по нестабильной связи, что приводит к её разрушению. В ходе этой реакции образуется 6-фосфоглюконат. Третья реакция. Катализирует реакцию 6-фосфоглюконатдегидрогеназа, а её кофермент — НАДФ. В этой реакции происходят два важных момента: глюконат теряет карбоксильную группу, и от пентозы отщепляются два водорода. Продукты реакции — рибулозо-5-фосфат и НАДФH.
Второй этап – этап структурных перестроек, Молекулы рибулозо-5- фосфата изомеризуются до рибозо-5- фосфата и ксилулозо-5- фосфата. Конечные продукты реакций : фруктозо-6- фосфат и глицеральдегидфосфат ( реакции гликолиза) (пентозофосфатный путь называют шунт , или обходной путь окисления глюкозы). Глицеральдегид-3-фосфат в зависимости от условий и вида клеток может уйти во 2-й этап гликолиза.
Значение пентозофосфатного пути
Значение первого этапа пентозофосфатного пути заключается в синтезе НАДФН и рибозо-5-фосфата.
Образованный НАДФН используется:
для синтеза жирных кислот
1.холестерола и других стероидов,
2.для синтеза глутаминовой кислоты из α-кетоглутаровой кислоты (реакция восстановительного аминирования),
3.для синтеза дезоксирибонуклеотидов,
4.для систем защиты клетки от свободно-радикального окисления (антиоксидантная защита).
Рибозо-5-фосфат абсолютно необходим для синтеза пуриновых и пиримидиновых нуклеотидов и нуклеиновых кислот.
При определенных условиях клетка может подключать второй неокислительный этап. Благодаря этому рибозо-5-фосфат не будет накапливаться и его углероды перейдут в состав фруктозо-6-фосфата (например, в эритроците) и уйдут в гликолиз. Либо глицеральдегид-3-фосфат восстановится до глицерол- 3-фосфата и будет использован для синтеза триацилглицеролов.
65. Энергетический выход окисления одной молекулы глюкозы при гликолизе, аэробном окислении и прямом окислении. Регуляция углеводного обмена
При гликолизе 1 молекулы глюкозы образуются 2 молекулы пирувата и 2 молекулы АТФ.
В аэробных условиях глюкоза окисляется до СО2 и Н2О. Суммарное уравнение:
С6Н12О6 + 6О2 → 6СО2+ 6Н2О + 2880 кДж/моль.
Этот процесс включает несколько стадий:
1.Аэробный гликолиз. В нем происходит окисления 1 глюкозы до 2 ПВК, с образованием 2 АТФ (сначала 2 АТФ затрачиваются, затем 4 образуются) и 2 НАДН2;
2.Превращение 2 ПВК в 2 ацетил-КоА с выделением 2 СО2 и образованием 2 НАДН2;
3.ЦТК. В нем происходит окисление 2 ацетил-КоА с выделением 4 СО2, образованием 2 ГТФ (дают 2 АТФ), 6 НАДН2 и 2 ФАДН2;
4.Цепь окислительного фосфорилирования. В ней происходит окисления 10 (8) НАДН2, 2 (4) ФАДН2 с участием 6 О2, при этом выделяется 6 Н2О и синтезируется 34 (32) АТФ.
+В результате аэробного окисления глюкозы образуется 38 (36) АТФ, из них: 4 АТФ в реакциях субстратного фосфорилирования, 34 (32) АТФ в реакциях окислительного фосфорилирования. КПД аэробного окисления составит 65%.
Прямое окисление (пфп)

Регуляция
К гормонам, которые влияют на углеводный обмен, принадлежат пептиды инсулин и глюкагон, глюкокортикоид кортизол и катехоламин адреналин. Инсулин индуцирует синтез de novo гликоген-синтазы, а также некоторых ферментов гликолиза. Одновременно инсулин подавляет синтез ключевых ферментов глюконеогенеза (репрессия). Глюкагон как антагонист инсулина действует в противоположном направлении: индуцирует ферменты глюконеогенеза и репрессирует пируваткиназу, ключевой фермент гликолиза. Другие аффекты глюкагона основаны на взаимопревращении ферментов и опосредованы вторичным мессенджером цАМФ. По этому механизму тормозится синтез гликогена и активируется расщепление гликогена. Подобным образом действует и адреналин. Торможение пируваткиназы глюкагоном также обусловлено взаимопревращением ферментов.
Глюкокортикоиды, прежде всего кортизол, индуцируют все ключевые ферменты глюконеогенеза. Одновременно они индуцируют ферменты деградации аминокислот и обеспечивают тем самым глюконеогенез исходными соединениями.
Метаболиты. Высокие концентрации АТФ (АТР) и цитрата тормозят гликолиз путем аллостерической регуляции фосфофруктокиназы. Кроме того, АТФ тормозит пируваткиназу. Ингибитором пируваткиназы является ацетил-КоА. Все эти метаболиты образуются при распаде глюкозы (торможение конечным продуктом). АМФ (AMP), сигнал дефицита АТФ, активирует расщепление гликогена и тормозит глюконеогенез.
Б. Фруктозо-2,6-дифосфат
Важную роль в обмене веществ в печени играет фруктозо-2,6-дифосфат. Это сигнальное вещество образуется в незначительных количествах из фруктозо-6-фосфата и выполняет чисто регуляторную функцию: стимулирует гликолиз путем активации фосфофруктокиназы и подавляет глюконеогенез с помощью торможения фруктозо-1,5-дифосфатазы.
Образование и распад фруктозо-2,6-дифосфата катализируются одним и тем же белком. В нефосфорилированной форме этот белок вызывает образование фруктозо-2,6-дифосфата . После фосфорилирования цАМФ-зависимой киназой он действует как фосфатаза и катализирует превращение фруктозо-2,6-дифосфата в фруктозо-6-фосфат. В присутствии адреналина и глюкагона в клетках печени повышается уровень цАМФ, т.е. оба гормона воздействуют как на гликолиз, так и на глюконеогенез. Суммарным результатом является быстрое повышение уровня глюкозы в крови.
66. Переваривание и всасывание белков в жкт.
Механизм действия протеолитических ферментов.Гниение белков в кишечнике под влиянием микроорганизмов.Обезвреживание продуктов гниения.
В пищевых продуктах содержание свободных аминокислот очень мало. Подавляющее их количество входит в состав белков, которые гидролизуются в ЖКТ под действием ферментов протеаз. Субстратная специфичность этих ферментов заключается в том, что каждый из них с наибольшей скоростью расщепляет пептидные связи, образованные определёнными аминокислотами. Протеазы, гидролизующие пептидные связи внутри белковой молекулы, относят к группе эндопептидаз. Ферменты, относящиеся к группе экзопептидаз, гидролизуют пептидную связь, образованную концевыми аминокислотами. Под действием всех протеаз ЖКТ белки пищи распадаются на отдельные аминокислоты, которые затем поступают в клетки тканей. В ротовой полости переваривания и всасывания белков не происходит.Желудочный сок - продукт нескольких типов клеток. Обкладочные (париетальные) клетки стенок желудка образуют соляную кислоту, главные клетки секретируют пепсиноген. Добавочные и другие клетки эпителия желудка выделяют муцинсодержащую слизь. Париетальные клетки секретируют в полость желудка также гликопротеин, который называют "внутренним фактором" (фактором Касла). Этот белок связывает "внешний фактор" - витамин В12, предотвращает его разрушение и способствует всасыванию. В тонком кишечнике полипептиды подвергаются дальнейшему расщеплению проте- азами, имеющимися в соке поджелудочной железы и на поверхности микроворсинок энтероцитов. Различные панкреатические ферменты атакуют белковую молекулу в разных участках. Ферменты, гидролизирующие белки, подразделяют на эндопептидазы (трипсин, химотрипсин, эластаза) и экзопептидазы (карбопептидазы А и В). Эндопептидазы гидролизуют внутренние связи пептидов, а экзопептидазы отщепляют только концевые группы преимущественно нейтральных и основных аминокислот. В тонком отделе кишечника происходит дальнейший гидролиз пептидов до аминокислот. Туда поступает панкреатический сок,который содержит неактивные предшественники протеаз: трипсиноген, химотрипсиноген, прокарбоксипептидазу, проэластазу. Слизистой кишечника вырабатывается фермент энтеропептидаза, который активирует трипсиноген до трипсина, а последний уже все остальные ферменты. Протеолитические ферменты содержатся также в клетках слизистой кишечника, поэтому гидролиз небольших пептидов происходит после их всасывания. Конечный результат действия ферментов желудка и кишечника – расщепление почти всей массы пищевых белков до свободных аминокислот. Протеолиз — процесс ферментативного гидролиза белков, катализирующийся протеолитическими ферментами (протеазами). Действие протеолитических ферментов может быть разделено на две различные категории:
-ограниченный протеолиз, в котором протеаза специфически расщепляет одну или несколько пептидных связей в белке-мишени, что обычно приводит к изменению функционального состояния последнего: ферменты, например, при этом становятся активными, а прогормоны превращаются в гормоны;
-неограниченный или тотальный протеолиз, при котором белки распадаются до своих аминокислот.
Образование токсичных продуктов из аминокислот под действием микроорганизмов кишечника называют гниением белков в кишечнике. Ферменты бактерий расщепляют аминокислоты и превращают их в амины, фенолы, индол, скатол, сероводород и другие ядовитые для организма соединения. Под действием ферментов бактерий из аминокислоты тирозина могут образовываться фенол и крезол путём разрушения боковых цепей аминокислот микробами. Эти вещества всасываются клетками кишечника, транспортируются кровью по воротной вене в печень, где обезвреживание фенола и крезола может происходить путём конъюгации с остатком серной или с глюкуроновой кислотой в составе УДФ-глюкуроната. Конъюгация глюкуроновых кислот с фенолом и крезолом происходит при участии фермента УДФ-глюкуронилтрансферазы. Продукты конъюгации хорошо растворимы в воде и выводятся с мочой через почки. В кишечнике из аминокислоты триптофана микроорганизмы образуют индол и скатол. Бактерии разрушают боковую цепь триптофана, оставляя нетронутой кольцевую структуру.Индол образуется в результате отщепления бактериями боковой цепи, в виде серина или аланина. Скатол и индол обезвреживаются в печени в 2 этапа. Сначала в результате микросомального окисления они приобретают гидроксильную группу. Так, индол переходит в индоксил, а затем вступает в реакцию конъюгации с ФАФС, образуя индоксилсерную кислоту, калиевая соль которой получила название животного индикана. Синтез гиппуровой кислоты из бензойной кислоты и глицина протекает у человека и большинства животных преимущественно в печени.
.Значение белков в питании.Баланс азота,азотистое равновесие,заменимые и незаменимые аминокислоты.
+Белки выполняют функцию основного строительного материала для роста и обновления клеток. Они входят в состав гормонов, ферментов и антител. Важнейшей функцией пищевых белков является обеспечение организма пластическим материалом. Основным источником, восполняющим фонд аминокислот организма, являются белки пищи. Аминокислоты (свободные и в составе белков) содержат почти 95% всего азота, поэтому баланс азота в организме является показателем состояния белкового и аминокислотного обмена. Азотистый баланс определяют как разницу между количеством азота, поступающего с пищей, и количеством выделяемого азота (преимущественно в виде мочевины и аммонийных солей). Азотистый баланс может быть равным нулю (азотистое равновесие)- у здорового человека при сбалансированном питании количество поступающего азота в сутки равно количеству выделяемого.Незмаменимые аминокислоты- те аминокислоты, синтез которых сложен и неэкономичен для организма, которые выгоднее получать с пищей. К ним относят фенилаланин, метионин, треонин, триптофан, валин, лизин, лейцин, изолейцин. Две аминокислоты - аргинин и гистидин - у взрослых образуются в достаточных количествах, однако детям для нормального роста организма необходимо дополнительное поступление этих аминокислот с пищей. Поэтому их называют частично заменимыми. Две другие аминокислоты - тирозин и цистеин - условно заменимые, так как для их синтеза необходимы незаменимые аминокислоты. Тирозин синтезируется из фенилаланина, а для образования цистеина необходим атом серы метионина. Остальные аминокислоты легко синтезируются в клетках и называются заменимыми. К ним относят глицин, аспарагиновую кислоту, аспарагин, глутаминовую кислоту, глутамин, серии, пролин, аланин. Если белок содержит все незаменимые аминокислоты в необходимых пропорциях и легко подвергается действию протеаз, то биологическая ценность такого белка условно принимается за 100, и он считается полноценным. К таким относят белки яиц и молока.