

56. Окисление ацетил-КоА в цикле Кребса. Связь между общими путями катаболизма и цепью переноса электронов и протонов. Цикл трикарбоновых кислот
Цикл протекает в матриксе митохондрий и представляет собой окисление молекулы ацетил-SКоА в восьми последовательных реакциях.
В первой реакции связываются ацетил и оксалоацетат (щавелевоуксусная кислота) с образованием цитрата (лимонной кислоты), далее происходит изомеризация лимонной кислоты до изоцитрата и две реакции дегидрирования с сопутствующим выделением СО2 и восстановлением НАД(образуются НАДН и FADH2).
В пятой реакции образуется ГТФ, это реакция субстратного фосфорилирования. Далее последовательно происходит ФАД-зависимое дегидрирование сукцината (янтарной кислоты), гидратация фумаровой кислоты до малата (яблочная кислота), далее НАД-зависимое дегидрирование с образованием оксалоацетата.
В итоге после восьми реакций цикла вновь образуется оксалоацетат.
Энергетический выход – 12АТФ.
Взаимосвязь цикла Кребса и дыхательной цепи: происходит через НАДН, который является донором для дыхательной цепи.
Последовательность реакций цитратного цикла
Образование цитрата
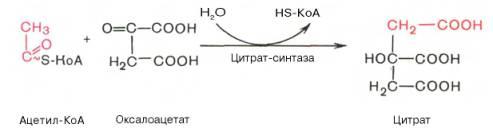
В реакции образования цитрата углеродный атом метильной труппы ацетил-КоА связывается с карбонильной группой оксалоацетата; одновременно расщепляется тиоэфирная связь и освобождается коэнзим A. Реакция сопровождается потерей большого количества энергии в виде теплоты. Катализирует реакцию цитрат-синтаза, фермент, локализованный в матриксе митохондрий.
Превращение цитрата в изоцитрат
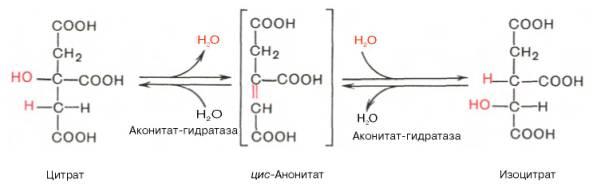
Вторая реакция цитратного цикла - обратимое превращение цитрата в изоцитрат (рис. 6-24). Фермент, катализирующий эту реакцию, назван аконитазой по промежуточному продукту, цис-аконитовой кислоте, которая предположительно образуется в реакции. Однако это соединение не обнаруживается в свободном виде, так как не отделяется от активного центра фермента до завершения реакции.
Окислительное декарбоксилирование изоцитрата
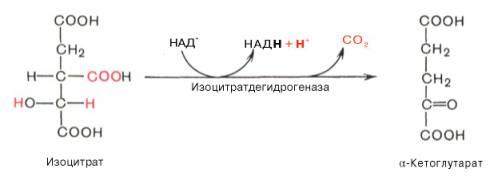
Эту реакцию катализирует изоцитратдегидрогеназа. В результате действия этого фермента на изоцитрат образуется α-кетоглутарат Реакция, катализируемая NAD-зависимой изоцитратдегидрогеназой, - самая медленная реакция цитратного цикла. АДФ - аллостерический активатор фермента.
Окислительное декарбоксилирование α-кетоглутарата
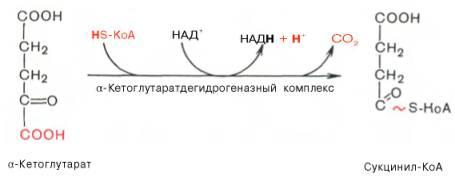
В этой реакции α-кетоглутарат подвергается окислительному декарбоксилированию с образованием в качестве конечных продуктов сукцинил-КоА, СО2 и NADH + Н+. В результате этой реакции образуется сукцинил-КоА Реакцию катализирует α-кетоглутаратдегидрогеназный комплекс.
Превращение сукцинил-КоА в сукцинат
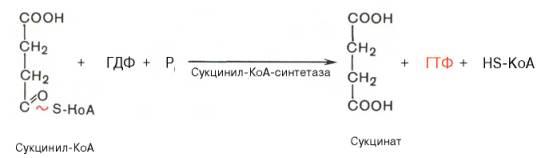
Сукцинил-КоА - высокоэнергетическое соединение. Изменение свободной энергии гидролиза этого тиоэфира составляет ΔG0'= -35,7 кДж/моль. В митохондриях разрыв тиоэфирной связи сукцинил-КоА сопряжён с реакцией фосфорилирования гуанозиндифосфата (ГДФ) до гуанозинтрифосфата (ГТФ).
Сукцинил-КоА → Сукцинат (ΔG0 = -10,36 кДж/моль).
Эту сопряжённую реакцию катализирует сукцинаттиокиназа. Промежуточный этап реакции - фосфорилирование молекулы фермента по одному из гистидиновых остатков активного центра. Затем остаток фосфорной кислоты присоединяется к ГДФ с образованием ГТФ. С ГТФ концевая фосфатная группа может переноситься на АДФ с образованием АТФ; эту обратимую реакцию катализирует нуклеозид-дифосфаткиназа.
ГТФ + АДФ ↔ ГДФ + АТФ.
Образование высокоэнергетической фосфо-ангидридной связи за счёт энергии субстрата (сукцинил-КоА) - пример субстратного фосфорилирования.