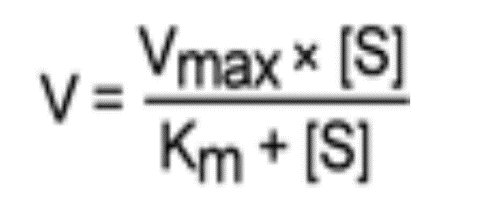20. Особенности ферментативного катализа. Специфичность действия ферментов. Особенности ферментативного катализа.
1. Белковая природа катализатора
2. Исключительно высокая эффективность. Эффективность биологического катализа превышает эффективность неорганического в 109 - 1012
3. Исключительно высокая специфичность:
а) абсолютная, когда фермент работает только со своим субстратом (фумараза с транс-изомерами фумаровой кислоты и не будет с цис-изомерами);
б) групповая - специфичен для узкой группы родственнных субстратов (ферменты ЖКТ).
4. Работает в мягких условиях (t=37, рН 7.0, определенные осмолярность и солевой состав).
5. Многоуровневая регуляция: регуляция активности на уровне условий среды, на уровне метаболона, на генетическом уровне, тканевом, клеточном, с помощью гормонов и медиаторов, а также с помощью субстратов и продуктов той реакции, которую они катализируют.
6. Кооперативность: ферменты способны организовывать ассоциации - продукт 1-го фермента, является субстратом для 2-го; продукт 2-го - субстратом для 3-го и т.д.
Кроме того, ферменты обладают адаптивностью, т. е. могут изменять свою активность и образовывать новые ассоциации.
7. Способны катализировать как прямую так и обратную реакцию. Направление реакции для многих ферментов определяется соотношением действующих масс.
8. Катализ жестко расписан, т. е. происходит поэтапно.
Специфичность, т.е. высокая избирательность действия ферментов, основана на комплементарности структуры субстрата и активного центра фермента.
1. Стереоспецифичность – катализ только одного из стереоизомеров, например:
• специфичность к L- или D-аминокислотам – например, почти все ферменты человека взаимодействуют с L-аминокислотами,
• специфичность к цис- и транс-изомерам. Например, аспартаза реагирует только с транс-изомером – фумаровой кислотой, но не с малеиновой кислотой (цис-изомер).
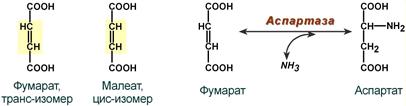
Стереоспецифичность аспартазы к транс-изомеру субстрата
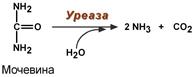
2. Абсолютная специфичность – фермент производит катализ только одного вещества. Например, каталаза разрушает перекись водорода, аргиназа расщепляет только аргинин, уреаза расщепляет только мочевину, глюкокиназа фосфорилирует только D-глюкозу.
3. Относительная групповая специфичность – превращение субстратов с некоторыми общими признаками. Например, цитохром Р450 окисляет только гидрофобные вещества, которых насчитывается около 7000. (альфа-амилаза – крахмал и его производные)
21. Кинетика ферментативных реакций. Факторы, определяющие скорость ферментативных реакций. Кинетика ферментативных реакций –
это раздел энзимологии, изучающий зависимость скорости химических реакций, катализируемых ферментами, от химической природы реагирующих веществ и факторов окружающей среды.
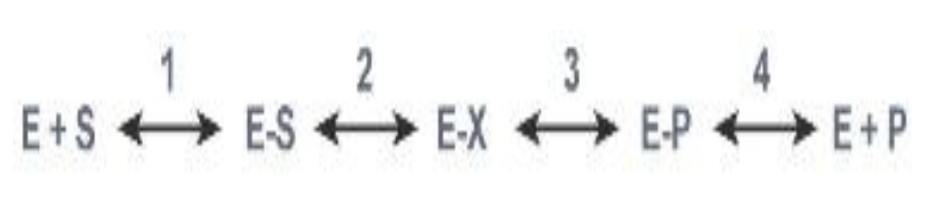
Этапы катализа
1. Присоединение субстрата (S) к ферменту (E) с образованием фермент-субстратного комплекса (E-S).
2. Преобразование фермент-субстратного комплекса в один или несколько переходных комплексов (E-X) за одну или несколько стадий.
3. Превращение переходного комплекса в комплекс фермент-продукт (E-P).
4. Отделение конечных продуктов от фермента.
Ферментативная активность зависит от: концентрации фермента, концентрации субстрата, концентрации кофактора, температуры, РН, присутствия ингибиторов
Для измерения каталитической активности ферментов используют такие показатели, как скорость реакции или активность фермента. Скорость ферментативной реакции определяется уменьшением количества молекул субстрата или увеличением количества молекул продукта за единицу времени. Скорость ферментативной реакции является мерой каталитической активности фермента.
На практике пользуются условными величинами, характеризующими активность фермента: 1 международная единица активности ( 1МЕ) соответствует такому количеству фермента, которое катализирует превращение 1 мкмоль субстрата за 1 минуту при оптимальных условиях (температура 37°С, оптимальное значение рН раствора). Эти единицы используют в медицинской и фармацевтической практике для оценки активности ферментов:
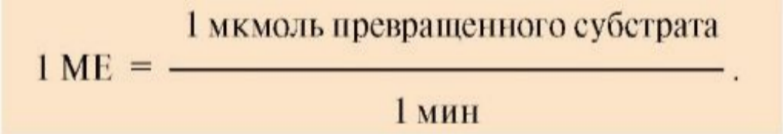
В системе СИ единица активности фермента – 1 Катал (Кат) –это количество фермента, катализирующее превращение 1 Моля субстрата за 1 секунду. (1 кат = моль / с)
Для оценки количества молекул фермента среди других белков данной ткани определяют удельную активность (Уд.Ак.) фермента, численно равную количеству превращенного субстрата (в мкмолях) за единицу времени одним миллиграммом (мг) белка (фермента, выделенного из ткани). По удельной активности судят о степени очистки фермента: чем меньше посторонних белков, тем выше удельная активность.

Кинетику исследуют в оптимальных условиях проведения энзиматической реакции. Оптимальные условия индивидуальны для каждого фермента и определяются в первую очередь температурой, при которой проводится реакция, и значением рН раствора.
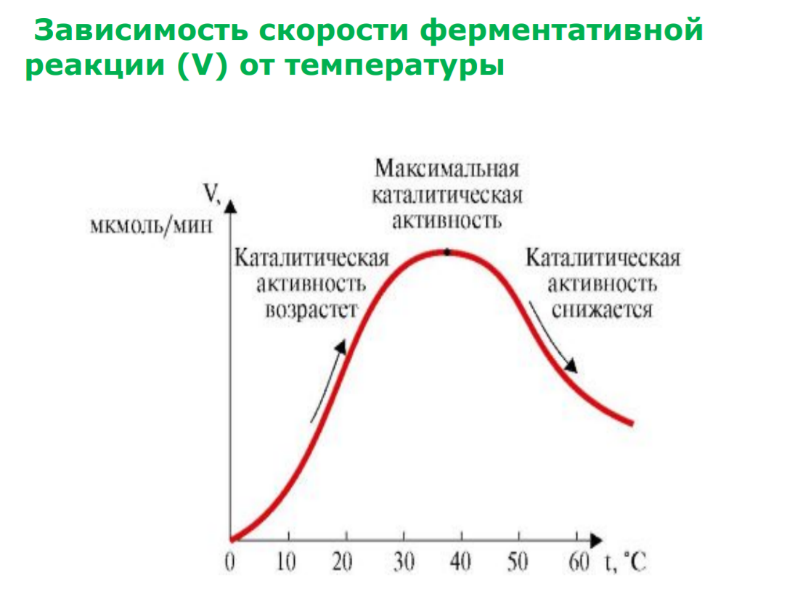
Зависимость скорости ферментативной реакции (V) от температуры . При увеличении температуры до + 40 С скорость ферментативной реакции возрастает. Выше +40 С скорость замедляется, т. к. начинается денатурация фермента. Возрастание скорости реакции при повышении температуры используется организмом как защитный механизм.
Понижение температуры (гипотермия организма и его отдельных тканей) используется в медицине для замедления ферментативных реакций при выполнении хирургических операций, при консервировании тканей и органов. При повышении t на 1 градус активность ферментов возрастает на 10%.
(Правило Вант-Гофа: при повышении t на каждые 10 градусов скорость реакции увеличивается примерно вдвое). Правило Вант-Гофа для ферментативных реакций справедливо лишь до +50-60 С.
Зависимость скорости ферментативной реакции (V) от рН среды. Для каждого фермента существует оптимальное значение рН, при котором наблюдается его максимальная активность. Оптимум РН для многих ферментов лежит в нейтральной среде. Исключения : пепсин РН = 1,5-2, щелочная фосфатаза (ЩФ) РН= 9-10.
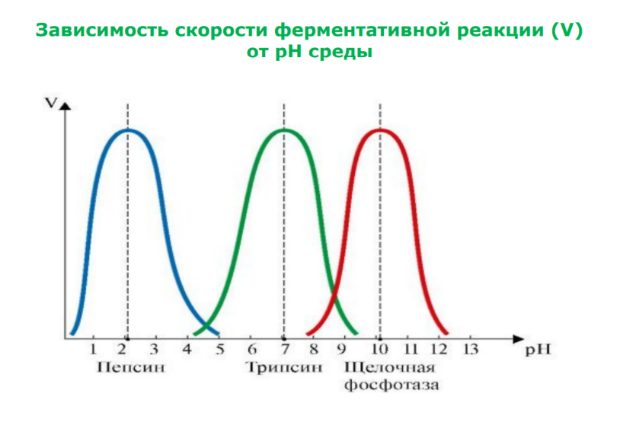
Изменение РН в организме при патологии нарушает функцию ферментов (изменяется степень ионизации функциональных групп активного и аллостерического центров и их архитектоника).
Влияние количества фермента: Зависимость прямо пропорциональна – чем больше фермента в клетке, тем выше скорость реакции. Количество фермента в клетке зависит от её потребности в ферменте.
В
покое в клетке функционирует лишь
небольшая часть ферментов. В ситуации
стресса в кинетике химической реакции
участвует максимальное количество
ферментов, что обеспечивает адаптацию
клетки к экстремальным условиям. 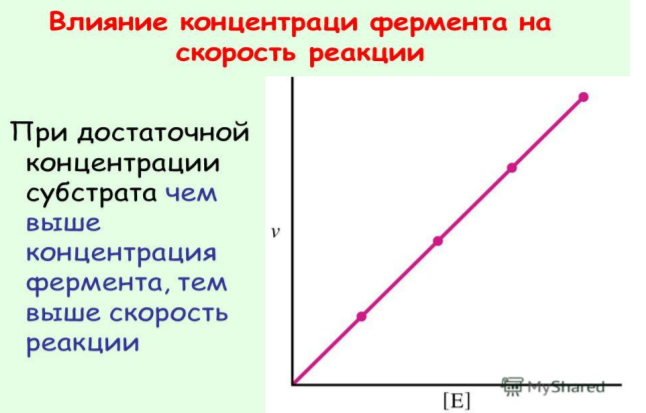
Скорость ферментативной реакции измеряют сразу после начала реакции (начальная скорость). График зависимости скорости ферментативной реакции от концентрации субстрата имеет вид гиперболы (насыщение миоглобина кислородом).
Основной кинетической характеристикой эффективности фермента является константа Михаэлиса - Km . Km – это концентрация субстрата, при которой скорость химической реакции равна 1/2 от максимально возможной. Km -величина постоянная, характеризует сродство данного фермента к субстрату .
Зависимость скорости ферментативных реакций от концентрации субстрата: чем меньше Km , тем больше сродство фермента к данному субстрату, тем выше начальная скорость реакции.
В математическом смысле Km соответствует концентрации субстрата при которой скорость реакции равна половине максимальной. Ее биологический смысл заключается в характеристике сродства фермента к субстрату: увеличение величины Кm означает снижение сродства фермента к субстрату.