

19. Общие пути обмена аминокислот. Биосинтез аминокислот.
 Общие
пути обмена аминокислот включают реакции
дезаминирования, трансаминирования
и декарбоксилирования:
Общие
пути обмена аминокислот включают реакции
дезаминирования, трансаминирования
и декарбоксилирования:
Реакции трансаминирования (переаминирования) - наиболее важные реакции среди общих путей обмена аминокислот.
Они играют основную роль в процессе глюконеогенеза и образования новых аминокислот.
Суть этой реакции - обратимый перенос аминогруппы с аминокислоты на кетокислоту без промежуточного образования аммиака.
Катализируют реакции трансаминирования ферменты трансаминазы (аминотрансферазы), в роли кофермента выступает пи- ридоксальфосфат (активная форма витамина В6):
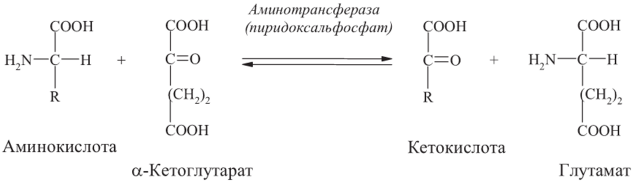
В переаминировании участвует много аминокислот (кроме лизина, треонина), наиболее активными являются глутаминовая, аспарагиновая кислоты, что связывают с высоким содержанием в тканях животных двух специфических аминотрансфераз - аспартатаминотрансферазы и аланинаминотрансферазы.
Аминотрансферазы участвуют в синтезе новых аминокислот из а-кетоглутарата.
Этот синтез возможен и для получения незаменимых аминокислот, если с пищей будут поступать соответствующие кетокислоты.
Дезаминирование - реакция, в ходе которой аминогруппа освобождается в виде аммиака.
Механизмы дезаминирования могут быть различными:
Восстановительное дезаминирование (с образованием насыщенной жирной кислоты):

Гидролитическое дезаминирование (с образованием карбоновой гидрокси- кислоты):

Элиминирующее дезаминирование (с образованием ненасыщенных жирных кислот):
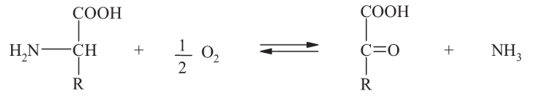
Окислительное дезаминирование (с образованием кетокислот). У человека окислительное дезаминирование является основным путем катаболизма аминокислот:
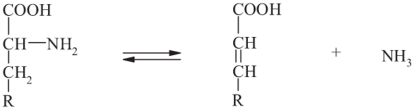
Выделяют два типа окислительного дезаминирования:
прямое
непрямое.
Если в ходе дезаминирования аминогруппа сразу превращается в аммиак, то реакция называется прямым окислительным дезаминированием. Прямое окислительное дезаминирование может происходить как в аэробных, так и в анаэробных условиях:
1 .
Аэробное прямое окислительное
дезаминирование - катализируется
оксидазами D- и
L-аминокислот с участием коферментов
ФАД и ФМН соответственно. Реакции,
катализируемые оксидазами, в клетках
протекают медленно.
.
Аэробное прямое окислительное
дезаминирование - катализируется
оксидазами D- и
L-аминокислот с участием коферментов
ФАД и ФМН соответственно. Реакции,
катализируемые оксидазами, в клетках
протекают медленно.
Анаэробное прямое окислительное дезаминирование - существует только для L-глутаминовой кислоты и катализируется ферментом глутаматдегидрогеназой.
Фермент присутствует в митохондриях всех клеток организма (за исключением мышечных). Глутаматдегидрогеназная реакция нуждается в присутствии коферментов НАД+ или НАДФ+:
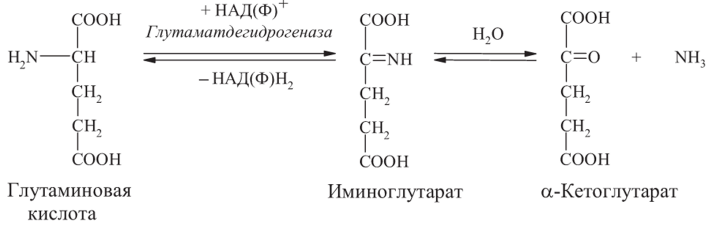
В ходе дезаминирования глутамата аминогруппа сразу превращается в аммиак.
Реакция дезаминирования глутамата ферментом глутаматдегидрогеназой обратима.
Обратная реакция - восстановительное аминирование а-кетоглутарата - служит источником глутамата в клетках и обезвреживает аммиак.
Все аминокислоты, кроме глутаминовой, дезаминируются непрямым путем. Непрямое окислительное дезаминирование активно протекает во всех клетках организма и состоит из двух этапов:
Трансаминирования с а-кетоглутаратом с образованием глутамата;
Прямого окислительного дезаминирования глутамата.
В результате трансаминирования а-кетоглутарат превращается в глутамат, а аминокислоты теряют аминогруппы и превращаются в а-кетокислоты. Далее их углеродный скелет катаболизируется специфическими путями и вовлекается в цикл Кребса, где сгорает до углекислого газа и воды.
Глутаминовая кислота является коллектором всех аминогрупп, и только она подвергается прямому окислительному дезаминированию с образованием аммиака и а-кетоглутаровой кислоты.

Таким образом, реакция прямого окислительного дезаминирования тесно связана с трансаминированием аминокислот. Сопряжение реакций трансаминирования и дезаминирования направляет поток лишнего аминного азота в печень для синтеза мочевины и в почки для синтеза аммонийных солей.
Декарбоксилирование аминокислот катализируется декарбоксилазами, ко- ферментом которых является пиридоксальфосфат. Продукты декарбоксилирования обладают высокой биологической активностью, и с этим связано их название «Биогенные амины»
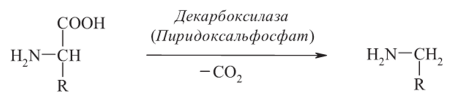
Биосинтез аминокислот:
Растения и многие виды бактерий содержат ферменты, необходимые для синтеза всех необходимых -кетокислот.
Животные утратили способность синтезировать некоторые -кетокислоты, из которых могут быть получены незаменимые аминокислоты.
Другие -кетокислоты могут образовываться в результате метаболизма иных соединений (в основном из глюкозы) и использоваться для синтеза заменимых аминокислот.
Человек и животные способны синтезировать из неаминокислотных предшественников только 8 из 20 аминокислот, необходимых для синтеза белка (заменимые аминокислоты), 2 аминокислоты (частично заменимые аминокислоты) синтезируются в ограниченных количествах и еще 2 образуются из незаменимых аминокислот (условно заменимые аминокислоты).
Углеродный скелет аминокислот образуется из промежуточных метаболитов гликолиза, пентозофосфатного пути, цикла Кребса.
Основными путями биосинтеза аминокислот являются:
прямое аминирование -кетокислот или ненасыщенных органических кислот,
реакции трансаминирования,
ферментативные взаимопревращения отдельных аминокислот – как заменимых, так и незаменимых.
Ниже приведена схема синтеза одиннадцати заменимых аминокислот, углеродный скелет которых может образовываться из метаболитов гликолиза и цикла Кребса:
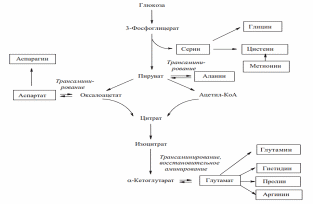
Необходимо отметить, что источником атома серы в молекуле условно заменимого цистеина служит незаменимый метионин. Двенадцатая аминокислота – тирозин – синтезируется путем гидроксилирования незаменимой аминокислоты фенилаланина и на схеме не показана.
Предшественниками для синтеза аминокислот служат промежуточные продукты метаболизма, такие кислоты, как:
α-кетоглутаровая,
щавелевоуксусная,
пировиноградная,
3-фосфоглицериновая и другие соединения.