

26. Биосинтез жирных кислот, химизм, ферменты.
Основные источники водорода для синтеза жирных кислот
В каждом цикле биосинтеза пальмитиновой кислоты проходят 2 реакции восстановления, донором водорода в которых служит кофермент NADPH.

Восстановление NADP+ происходит в реакциях:
дегидрирования в окислительных стадиях пентозофосфатного пути катаболизма глюкозы;
дегидрирования малата малик-ферментом;
дегидрирования изоцитрата цитозольной NADP-зависимой дегидрогеназой.
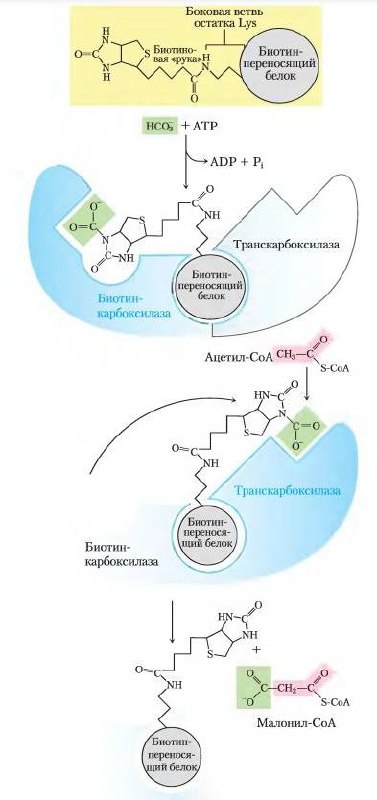 Рисунок
1 Реакция ацетил-СоА-карбоксилазы
Рисунок
1 Реакция ацетил-СоА-карбоксилазы
Синтез жирных кислот протекает в цитоплазме клетки. В митохондриях в основном происходит удлинение существующих цепей жирных кислот. Установлено, что в цитоплазме печеночных клеток синтезируется пальмитиновая кислота (16 углеродных атомов), а в митохондриях этих клеток из уже синтезированной в цитоплазме клетки пальмитиновой кислоты или из жирных кислот экзогенного происхождения, т.е. поступающих из кишечника, образуются жирные кислоты, содержащие 18, 20 и 22 углеродных атома.
Биосинтез жирных кислот протекает с участием НАДФН, АТФ, Мn2+ и НСО3 – (в качестве источника СО2); субстратом является ацетил-КоА, конечным продуктом – пальмитиновая кислота.
Происходит образование малонилапю, а затем начинается "нанизывание" атомов углерода на цепь, пока не получится пальмитиновая, либо другая жирная кислота.
Регуляция биосинтеза и окисления жирных кислот
Глюкагон и адреналин:
увеличивают скорость β-окисления,
снижают скорость синтеза жирных кислот
Инсулин:
снижает скорость β-окисления,
увеличивает скорость синтеза жирных кислот
27. Биосинтез жирных кислот с длинной цепью углеродных атомов и непредельных.
В настоящее время показано, что в микросомах клеток млекопитающих образование двойных связей может происходить только на участке цепи жирной кислоты от 9-го до 1-го углеродных атомов, ибо в микросомах отсутствуют десатуразы, которые могли бы катализировать образование двойных связей в цепи далее 9-го углеродного атома. У животных двойные связи могут образовываться в Δ4-, Δ5-, Δ6- и Δ9-положении, но не далее Δ9-положения, в то время как у растений – в Δ6-, Δ9-, Δ12 и Δ15-положении. Поэтому в организме млекопитающих, в том числе и человека, не могут образовываться, например, из стеариновой кислоты (18:0) линолевая (18:2; 9,12) и линоленовая (18:3; 9,12,15) кислоты. Эти кислоты относятся к категории незаменимых жирных кислот. К незаменимым жирным кислотам обычно относят также арахидоновую кислоту (20:4; 5,8,11,14).


Полиненасыщенными жирными кислотами принято называть одноосновные жирные кислоты, в структуре которых присутствуют две и более двойных связей между атомами углерода.
К полиненасыщенным жирным кислотам относятся в числе прочих и незаменимые жирные кислоты или эссенциальные жирные кислоты, получившие название витамина F, такие как линолевая (две двойные связи, положение первой – омега-6, то есть при шестом атоме углерода, отсчитывая от метильного конца) и линоленовая(три двойные связи, положение первой – омега-3, т. е. при третьем атоме углерода),эйкозапентаеновая (шесть двойных связей, положение первой – omega-3) идокозагексаеновая (пять двойных связей, положение первой – омега-3) кислоты.