

Metabolicheskaia_biokhimiia
.pdf181
Панкреатическая липаза (КФ 3.1.1.3) является гликопротеидом, имеющим мол. массу 48000 (у человека) и оптимум рН 8–9. Данный фермент расщепляет триглицериды, находящиеся в эмульгированном состоянии (действие фермента на растворенные субстраты значительно слабее). Как и другие пищеварительные ферменты (пепсин, трипсин, химотрипсин), панкреатическая липаза поступает в верхний отдел тонкой кишки в виде неактивной пролипазы. Превращение пролипазы в активную липазу происходит при участии желчных кислот и еще одного белка панкреатического сока – колипазы (мол. масса 10000). Последняя присоединяется к пролипазе в молекулярном соотношении 2:1. Это приводит к тому, что липаза становится активной и устойчивой к действию трипсина. Установлено, что основными продуктами расщепления триглицеридов при действии панкреатической липазы являются β(2)-моноглицерид и жирные кислоты. Фермент катализирует гидролиз эфирных связей в α(1), α(3)- положениях, в результате чего и образуются β(2)-моноглицерид и две частицы (молекулы) жирной кислоты. На скорость катализируемого липазой гидролиза триглицеридов не оказывает существенного влияния ни степень ненасыщенности жирных кислот, ни длина ее цепи (от С12 до С18). Гидролиз триглицеридов при участии панкреатической липазы можно изобразить в виде следующей схемы:
В панкреатическом соке наряду с липазой содержится моноглицеридная изомераза –фермент, катализирующий внутримолекулярный перенос ацила из β(2)-положения моноглицерида в
181
182
α(1)-положение. В процессе переваривания пищевых жиров при участии этого фермента примерно треть βмоноглицерида превращается в αмоноглицерид. Поскольку эфирная связь в αположении чувствительна к действию панкреатической липазы, последняя расщепляет большую часть αмоноглицеридов до конечных продуктов – глицерина и жирной кислоты. Меньшая часть αмоноглицеридов успевает всосаться в стенку тонкой кишки, минуя воздействие липазы.
Всасывание триглицеридов и продуктов их расщепления.
Всасывание происходит в проксимальной части тонкой кишки. Тонкоэмульгированные жиры (величина жировых капель эмульсии не должна превышать 0,5 мкм) частично могут всасываться через стенки кишечника без предварительного гидролиза. Основная часть жира всасывается лишь после расщепления его панкреатической липазой на жирные кислоты, моноглицериды и глицерин. Жирные кислоты с короткой углеродной цепью (менее 10 атомов углерода) и глицерин, будучи хорошо растворимыми в воде, свободно всасываются в кишечнике и поступают в кровь воротной вены, оттуда в печень, минуя какие-либо превращения в кишечной стенке. Более сложно происходит всасывание жирных кислот с длинной углеродной цепью и моноглицеридов. Этот процесс осуществляется при участии желчи и главным образом желчных кислот, входящих в ее состав. В желчи соли желчных кислот, фосфолипиды и холестерин содержатся в соотношении 12,5:2,5:1,0. Жирные кислоты с длинной цепью и моноглицериды в просвете кишечника образуют с этими соединениями устойчивые в водной среде мицеллы. Структура мицелл такова, что их гидрофобное ядро (жирные кислоты, моноглицериды и др.) оказывается окруженным снаружи гидрофильной оболочкой из желчных кислот и фосфолипидов. Мицеллы примерно в 100 раз меньше самых мелких эмульгированных жировых капель. В составе мицелл высшие жирные кислоты и моноглицериды переносятся от места гидролиза жиров к всасывающей поверхности кишечного эпителия. Относительно механизма всасывания жировых мицелл единого мнения нет. Одни исследователи считают, что в результате так называемой мицеллярной диффузии, а возможно, и пиноцитоза мицеллы целиком проникают в эпителиальные клетки ворсинок, где происходит распад жировых мицелл. При этом желчные кислоты сразу поступают в ток крови и через систему воротной вены попадают сначала в печень, а оттуда вновь в желчь. Другие исследователи допускают возможность перехода в клетки ворсинок только липидного компонента жировых мицелл. Соли желчных кислот, выполнив свою физиологическую роль, остаются в просвете кишечника; позже основная масса их всасывается в кровь (в подвздошной кишке), попадает в печень и затем выделяется с желчью. Таким образом, все исследователи признают, что происходит постоянная циркуляция желчных кислот между печенью и кишечником. Этот процесс получил название печеночно-кишечной (гепатоэнтеральной) циркуляции. С помощью метода меченых атомов было показано, что в желчи содержится лишь небольшая часть желчных кислот
182
183
(10–15% от общего количества), вновь синтезированных печенью. Таким образом, основная масса желчных кислот (85–90%) –это желчные кислоты, реабсорбированные в кишечнике и повторно секретируемые в составе желчи. Установлено, что у человека общий пул желчных кислот составляет примерно 2,8–,5 г, при этом они совершают 6– 8 оборотов в сутки.
Расщепление и всасывание фосфолипидов и холестерина. Подавляющая часть фосфолипидов содержимого тонкой кишки приходится на фосфатидилхолин (лецитин), основная масса которого поступает в кишечник с желчью (11–12 г/сут) и меньшая часть (1–2 г/сут) –с пищей. Существует две точки зрения относительно судьбы поступивших в тонкую кишку экзогенных и эндогенных фосфолипидов. Согласно одной из них, и те, и другие фосфолипиды подвергаются в кишечнике атаке со стороны фосфолипазы А2, катализирующей гидролиз сложноэфирной связи в βположении. В результате катализируемой фосфолипазой А2 реакции глицерофосфолипиды расщепляются с образованием лизофосфолипида и жирной кислоты. Лизофосфолипид может подвергаться расщеплению при действии другого фермента панкреатического сока – лизофосфолипазы. В результате из лизолецитина освобождается последняя частица жирной кислоты и образуется глицерофосфохолин, который хорошо растворяется в водной среде и всасывается из кишечника в кровь. Сторонники другой точки зрения считают, что фосфолипиды «желчного» (более точно печеночного) происхождения в отличие от пищевых фосфолипидов не подвергаются воздействию фосфолипазы А2. Следовательно, функция «желчных» фосфолипидов исключительно связана с гепатоэнтеральной циркуляцией желчи: с желчью они поступают в кишечник, с желчными кислотами участвуют в мицеллярной солюбилизации липидов и вместе с ними возвращаются в печень. Таким образом, существует как бы два пула фосфолипидов в кишечнике: «желчный», защищенный от действия фосфолипазы А2, и «пищевой», подверженный ее действию. Пока трудно объяснить причину существования двух пулов фосфолипидов и их различное отношение к действию фосфолипазы А2. В зависимости от пищи организм взрослого человека получает ежедневно 300–00 мг холестерина, содержащегося в пищевых продуктах частично в свободном (неэстерицифицированном) виде, частично в виде эфиров с жирными кислотами. Эфиры холестерина расщепляются на холестерин и жирные кислоты особым ферментом панкреатического и кишечного соков – гидролазой эфиров холестерина, или холестеролэстеразой (КФ 3.1.1.13). В тонкой кишке происходит всасывание холестерина, источником которого являются: – холестерин пищи (0,3–0,5 г/сут; у вегетарианцев значительно меньше); – холестерин желчи (ежедневно с желчью выделяется 1–2 г эндогенного неэстерифицированного холестерина); – холестерин, содержащийся в слущенном эпителии пищеварительного тракта и в кишечных соках (до 0,5 г/сут). В общей сложности в кишечник поступает 1,8–2,5 г эндогенного и экзогенного холестерина. Из этого количества около 0,5 г холестерина выделяется с фекалиями в виде восстановленного продукта
183
184
– копростерина и очень небольшая часть в виде окисленных продуктов – холестенона и др. И восстановление, и окисление холестерина происходят в толстой кишке под воздействием ферментов микробной флоры. Основная часть холестерина в неэстерифицированной форме подвергается всасыванию в тонкой кишке в составе смешанных жировых мицелл, состоящих из желчных кислот, жирных кислот, моноглицеридов, фосфолипидов и лизофосфолипидов.
Ресинтез липидов в кишечной стенке
Триглицериды. По современным представлениям, ресинтез триглицеридов происходит в эпителиальных клетках (энтероцитах слизистой оболочки ворсинок тонкой кишки) двумя путями. Первый путь –β- моноглицеридный. Долгое время этот путь считался единственным. Суть его состоит в том, что βмоноглицериды и жирные кислоты, проникающие в процессе всасывания в эпителиальные клетки кишечной стенки, задерживаются в гладком эндоплазматическом ретикулуме клеток. Здесь из жирных кислот образуется их активная форма – ацил-КоА и затем происходит ацилирование βмоноглицеридов с образованием сначала диглицеридов, а затем триглицеридов:
β-Моноглицерид + R–СО–KoA → Диглицерид + HS-KoA;
Диглицерид + R1–СО–KoA → Триглицерид + HS-KoA
Все реакции катализируются ферментным комплексом –триглицерид- синтетазой, включающим в себя ацил-КоА-синтетазу, моноглицеридацилтрансферазу и диглицеридацилтрансферазу.
Второй путь ресинтеза триглицеридов протекает в шероховатом эндоплазматическом ретикулуме эпителиальных клеток и включает следующие реакции:
1)образование активной формы жирной кислоты –ацил-КоА при участии ацил-КоА-синтетазы;
2)образование αглицерофосфата при участии глицеролкиназы;
3)превращение αглицерофосфата в фосфатидную кислоту при участии глицерофосфат-ацилтрансферазы;
4)превращение фосфатидной кислоты в диглицерид при участии фосфатидат-фосфогидролазы;
5)ацилирование диглицерида с образованием триглицерида при участии диглицеридацилтрансферазы.
Как видно, первая и последняя реакции повторяют аналогичные реакции βмоноглицеридного пути. Установлено, что αглицерофосфатный путь ресинтеза жиров (триглицеридов) приобретает значение, если в эпителиальные клетки слизистой оболочки тонкой кишки поступили преимущественно жирные кислоты. В случае, если в стенку кишки поступили жирные кислоты вместе с β-моноглицеридами, запускается β- моноглицеридный путь. Как правило, наличие в эпителиальных клетках
184
185
избытка β-моноглицеридов тормозит протекание α-глицерофосфатного пути.
Ресинтез фосфолипидов в кишечной стенке. В энтероцитах наряду с ресинтезом триглицеридов происходит также и ресинтез фосфолипидов. В образовании фосфатидилхолинов и фосфатидилэтаноламинов участвует ресинтезированный диглицерид, а в образовании фосфатидилинозитолов – ресинтезированная фосфатидная кислота. Участие этих субстратов в образовании фосфолипидов в стенке кишечника происходит по тем же закономерностям, что и в других тканях. Необходимо подчеркнуть, что в стенке кишечника синтезируются жиры, в значительной степени специфичные для данного вида животного и отличающиеся по своему строению от пищевого жира. В известной мере это обеспечивается тем, что в синтезе триглицеридов (а также фосфолипидов) в кишечной стенке принимают участие наряду с экзогенными и эндогенные жирные кислоты. Однако способность к осуществлению в стенке кишечника синтеза жира, специфичного для данного вида животного, все же ограничена. Показано, что при скармливании животному (например, собаке), особенно предварительно голодавшему, больших количеств чужеродного жира (например, льняного масла или верблюжьего жира) часть его обнаруживается в жировых тканях животного в неизмененном виде. Жировая ткань, скорее всего, является единственной тканью, где могут откладываться чужеродные жиры. Липиды, входящие в состав протоплазмы клеток других органов и тканей, отличаются высокой специфичностью, их состав и свойства мало зависят от пищевых жиров.
Образование хиломикронов и транспорт липидов.
Ресинтезированные в эпителиальных клетках кишечника триглицериды и фосфолипиды, а также поступивший в эти клетки из полости кишечника холестерин (здесь он может частично эстерифицироваться) соединяются с небольшим количеством белка и образуют относительно стабильные комплексные частицы – хиломикроны (ХМ). Последние содержат около 2% белка, 7% фосфолипидов, 8% холестерина и его эфиров и более 80% триглицеридов. Диаметр ХМ колеблется от 0,1 до 5 мкм. Благодаря большим размерам частиц ХМ не способны проникать из эндотелиальных клеток кишечника в кровеносные капилляры и диффундируют в лимфатическую систему кишечника, а из нее – в грудной лимфатический проток. Затем из грудного лимфатического протока ХМ попадают в кровяное русло, т.е. с их помощью осуществляется транспорт экзогенных триглицеридов, холестерина и частично фосфолипидов из кишечника через лимфатическую систему в кровь. Уже через 1– ч после приема пищи, содержащей жиры, наблюдается алиментарная гиперлипемия. Это физиологическое явление, характеризующееся в первую очередь повышением концентрации триглицеридов в крови и появлением в ней ХМ. Пик алиментарной гиперлипемии наблюдается через 4 ч после приема жирной пищи. Обычно через 10–2 ч после приема пищи содержание триглицеридов возвращается к нормальным величинам, а ХМ полностью исчезают из кровяного русла. Известно, что печень и жировая ткань играют наиболее существенную роль в
185
186
дальнейшей судьбе ХМ. Последние свободно диффундируют из плазмы крови в межклеточные пространства печени (синусоиды). Допускается, что гидролиз триглицеридов ХМ происходит как внутри печеночных клеток, так и на поверхности. ХМ не способны (из-за своих размеров) проникать в клетки жировой ткани. В связи с этим триглицериды ХМ подвергаются гидролизу на поверхности эндотелия капилляров жировой ткани при участии фермента липопротеидлипазы.
5.1.2 Пути окисления жирных кислот
Общее количество жировой ткани у взрослого мужчины со средней массой тела равно примерно 20 кг, а у тучных людей – на десятки килограммов больше. Жировая ткань, состоящая в основном из жировых клеток, или адипоцитов, распространена по всему организму: под кожей, в брюшной полости, образует жировые прослойки вокруг отдельных органов. Около 65% от массы жировой ткани приходится на долю отложенных в ней триглицеридов, что составляет приблизительно 95% от всех триглицеридов организма. Известно, что главным источником жирных кислот, используемых в качестве «топлива», служит резервный жир, содержащийся в жировой ткани. Принято считать, что триглицериды жировых депо выполняют в обмене липидов такую же роль, как гликоген в печени в обмене углеводов, а высшие жирные кислоты по своей энергетической роли напоминают глюкозу, которая образуется в процессе фосфоролиза гликогена. При физической работе и других состояниях организма, требующих повышенных энергозатрат, потребление триглицеридов жировой ткани как энергетического резерва увеличивается.
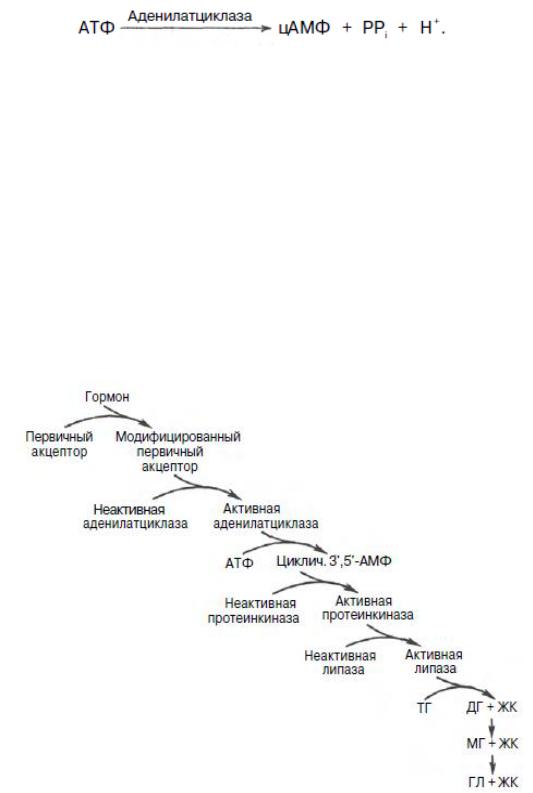
Липолиз триглицеридов в жировой ткани. В качестве источника энергии могут использоваться только свободные, т.е. неэстерифицированные, жирные кислоты. Поэтому триглицериды сначала гидролизуются при помощи специфических тканевых ферментов – липаз – до глицерина и свободных жирных кислот. Последние из жировых депо могут переходить в плазму крови (мобилизация высших жирных кислот), после чего они используются тканями и органами тела в качестве энергетического материала. В жировой ткани содержится несколько липаз, из которых наибольшее значение имеют триглицеридлипаза (так называемая гормоночувствительная липаза), диглицеридлипаза и моноглицеридлипаза. Активность двух последних ферментов в 1000 раз превышает активность первого. Триглицеридлипаза активируется рядом гормонов (например, адреналином, норадреналином, глюкагоном и др.), тогда как диглицеридлипаза и моноглицеридлипаза не чувствительны к их действию. Триглицеридлипаза является регуляторным ферментом. Установлено, что гормоночувствительная липаза (триглицеридлипаза) находится в жировой ткани в неактивной форме, и активация ее гормонами протекает сложным каскадным путем, включающим участие по крайней мере двух ферментативных систем. Процесс начинается со взаимодействия гормона с клеточным рецептором, в результате чего модифицируется структура
186
187
рецептора (сам гормон в клетку не поступает) и такой рецептор активирует аденилатциклазу (КФ 4.6.1.1). Последняя, как известно, катализирует образование циклического аденозинмонофосфата (цАМФ) из аденозинтрифосфата (АТФ):
Образовавшийся цАМФ активирует фермент протеинкиназу (КФ 2.7.1.37), который путем фосфорилирования неактивной триглицеридлипазы превращает ее в активную форму, рисунок 53. Активная триглицеридлипаза расщепляет триглицерид на диглицерид и жирную кислоту. Затем при действии ди- и моноглицеридлипаз образуются конечные продукты липолиза
– глицерин и свободные жирные кислоты, которые поступают в кровяное русло. Скорость липолиза триглицеридов не является постоянной, она подвержена регулирующему влиянию различных факторов, среди которых особое значение имеют нейрогормональные. Связанные с альбуминами плазмы крови в виде комплекса свободные жирные кислоты с током крови попадают в органы и ткани, где комплекс распадается, а жирные кислоты подвергаются либо βокислению, либо частично используются для синтеза триглицеридов, глицерофосфолипидов, сфингофосфолипидов и других соединений, а также на эстерификацию холестерина.
Рисунок 53. – Липолитический каскад (по Стайнбергу). ТГ - триглицериды; ДГ - диглицериды; МГ - моноглицериды;
ГЛ - глицерин; ЖК - жирные кислоты
187

188
Установлено, что окисление жирных кислот протекает в печени, почках, скелетных и сердечной мышцах, в жировой ткани. В мозговой ткани скорость окисления жирных кислот весьма незначительна; основным источником энергии в мозговой ткани служит глюкоза. В 1904 г. Ф. Кнооп (F. Knoop) выдвинул гипотезу βокисления жирных кислот на основании опытов по скармливанию собакам различных жирных кислот, в которых один атом водорода в концевой метильной группе (ω-углеродного атома) был замещен радикалом (С6Н5–). Ф. Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в βположении. В результате от молекулы жирной кислоты последовательно отщепляются двууглеродные фрагменты со стороны карбоксильной группы. Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют четное число углеродных атомов. Любая такая кислота, от которой отщепляется по паре углеродных атомов, в конце концов проходит через стадию масляной кислоты. После очередного βокисления масляная кислота становится ацетоуксусной. Последняя затем гидролизуется до двух молекул уксусной кислоты. Теория βокисления жирных кислот, предложенная Ф. Кноопом, в значительной мере послужила основой современных представлений о механизме окисления жирных кислот. Доставка жирных кислот к месту их окисления – к митохондриям – происходит сложным путем: при участии альбумина осуществляется транспорт жирной кислоты в клетку; при участии специальных белков (fatty acid binding proteins, FABP) – транспорт в пределах цитозоля; при участии карнитина – транспорт жирной кислоты из цитозоля в митохондрии. Процесс окисления жирных кислот складывается из следующих основных этапов.
Активация жирных кислот. Свободная жирная кислота независимо от длины углеводородной цепи является метаболически инертной и не может подвергаться никаким биохимическим превращениям, в том числе окислению, пока не будет активирована. Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима A (HS-KoA) и ионов Mg2+. Реакция катализируется ферментом ацил-КоА- синтетазой:
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты. Считают, что активация жирной кислоты протекает в 2 этапа. Сначала жирная кислота реагирует с АТФ с образованием ациладенилата, представляющим собой эфир жирной кислоты и АМФ. Далее сульфгидрильная группа КоА действует на прочно связанный с ферментом ациладенилат с образованием ацил-КоА и АМФ.
Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и
188

189
протекает их окисление. Переносчиком активированных жирных кислот с длинной цепью через внутреннюю митохондриальную мембрану служит карнитин.
Ацильная группа переносится с атома серы КоА на гидроксильную группу карнитина с образованием ацилкарнитина, который диффундирует через внутреннюю митохондриальную мембрану: Реакция протекает при участии специфического цитоплазматического фермента карнитинацилтрансферазы. Уже на той стороне мембраны, которая обращена к матриксу, ацильная группа переносится обратно на КоА, что термодинамически выгодно, поскольку О-ацильная связь в карнитине обладает высоким потенциалом переноса группы. Иными словами, после прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция –расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитин-ацилтрансферазы:
Внутримитохондриальное окисление жирных кислот. Процесс окисления жирной кислоты в митохондриях клетки включает несколько последовательных энзиматических реакций.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию, при этом ацил-КоА теряет 2 атома водорода в α и βположениях, превращаясь в КоА-эфир ненасыщенной кислоты. Таким образом, первой реакцией в каждом цикле распада ацил-КоА является его окисление ацил-КоА-дегидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3:
189

190
Существует несколько ФАД-содержащих ацил-КоА-дегидрогеназ, каждая из которых обладает специфичностью по отношению к ацил-КоА с определенной длиной углеродной цепи.
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА): Заметим, что гидратация еноил-КоА стереоспецифична, подобно гидратации фумарата и аконитата. В результате гидратации транс- 2-двойной связи образуется только L-изомер 3-гидроксиацил-КоА.
Вторая стадия дегидрирования. Образовавшийся βоксиацил-КоА (3-
гидроксиацил-КоА) затем дегидрируется. Эту реакцию катализируют НАД+- зависимые дегидрогеназы:
Тиолазная реакция. В ходе предыдущих реакций происходило окисление метиленовой группы при С-3 в оксогруппу. Тиолазная реакция представляет собой расщепление 3-оксоацил-КоА с помощью тиоловой группы второй молекулы КоА. В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетилКоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (βкетотиолазой):
Образовавшийся ацетил-КоА подвергается окислению в цикле
190