

Методы / Луизов Цвет и свет
.pdf5.2. Развитие трехкомпонентной теории.
Трехкомпонентная теория была развита Томасом Юнгом (1773–1829). Человек исключительно одаренный, Юнг был практикующим врачом в Лондоне и вместе с тем сделал выдающиеся открытия в различных областях науки: в механику ввел величину, которую называют модулем Юнга, в оптике, по существу, доказал волновую природу света своими опытами по дифракции, внес большой вклад в расшифровку египетских иероглифов. Если Ньютон показал, что разные цвета можно получать смешением других цветов, то Юнг ввел понятие о трех основных цветах и простым, изящным опытом показал, как их смешением можно получить другие цвета. Взяв три проекционных фонаря, Юнг направил их свет на белый экран так, чтобы проекции кругов частично перекрывались (рис. 5.2). В три фонаря были вставлены светофильтры: зеленый, красный и синий. В середине картины перекрывались изображения всех трех цветов и появлялся белый цвет. Перекрытие синего и зеленого давали голубой, зеленого и красного – желтый, красного и синего – пурпурный (45).
Юнгу было ясно, что трехкомпонентность цвета нельзя объяснить физическими свойствами самого света. Значит, она зависит от свойств глаза. Глаз имеет три приемника с различными чувствительностями к разным областям спектра. Суммирование степени их возбуждения светом дает ощущение того или иного цвета.
Рис. 5.2 Смешение цветов по Юнгу.
Д. К. Максвелл (1831–1879), создавший электромагнитную теорию света, очень интересовался цветовым зрением. Максвелл построил первый колориметр, в котором измеряемый цвет Ц освещал половину поля зрения, в то время как другая освещалась смесью трех излучений: красного, зеленого и синего. Интенсивности каждого из излучений можно было изменять до тех пор, пока не устанавливалось равенство цвета обоих полей
прибора. После установления равенства можно было написать уравнение |
|
Ц = аК + bЗ + сС, |
(5.1) |
где Ц – измеряемый цвет, а коэффициенты а, b и с – величины, пропорциональные интенсивностям красного (К), зеленого (3) и синего (С) излучения соответственно. У Максвелла эти коэффициенты были равны или пропорциональны ширине щели, пропускающей каждое из трех излучений к полю сравнения.
За основные Максвелл принял три спектральных цвета с длинами волн λк = 630 нм, λ3 = 528 нм и λс = 457 нм. Графически смешение цветов Максвелл изобразил в виде равностороннего треугольника (рис. 5.3). Стороны треугольника нанесены штрихами, в вершинах лежат излучения с λк λ3, λс. Цвет, получающийся в результате сложения интенсивностей, пропорциональных коэффициентам а, b и с для основных цветов, можно найти по принципу отыскания центра тяжести, как и в круге Ньютона. При равенстве а = b = с получается белый цвет (центр треугольника).
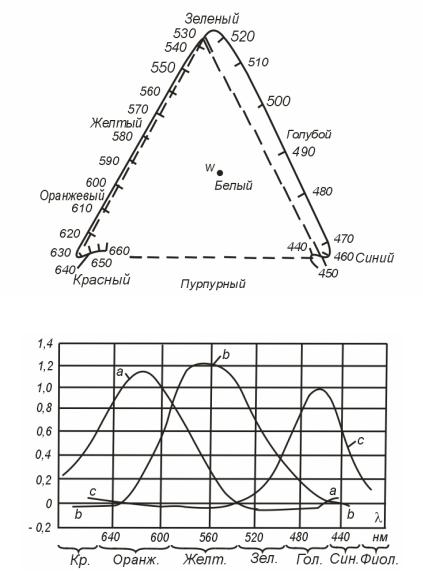
Рис. 5.3 Цветовой треуголник Максвелла.
Рис. 5.4 Значения коэффициентов Максвелла a, b и c, для спектрально-чистых цветов.
Спектрально-чистые цвета лежат на сплошной линии за пределами штрихового треугольника. При положительных значениях всех трех коэффицигытов а, b и с можно получить только цвета, лежащие внутри треугольника и на его сторонах.
Пусть по правилу сложения сил мы нашли для некоторого цвета Ц его положение на треугольнике Максвелла. Проведя из точки белого цвета W через любую точку Ц прямую до ее пересечения со сплошной линией спектрально-чистых тонов, мы получаем длину волны λ цвета Ц. Чем дальше точка Ц от точки W, тем больше чистота цвета Ц. Таким образом, точка на чертеже дает возможность судить о значениях величин λ и ρ для зтого цвета. Яркость L зависит от абсолютиых значений величин а, b и с.
На рис. 5.4 представлены в виде кривых найденные Максвеллом значения коэффициентов а, b и с для спектральных цветов. Смысл отрицательных коэффициентов мы поясним при описании совремеиной колориметрической системы RGB.
Крупный ученый Г. Гельмгольц (3821–1894), сделавший много открытий в различных областях физики, признал трехкомпонентную теорию зрения, способствовал ее развитию, построил свою модель колориметра. На колориметре Гельмгольца работали Кениг и Дитеричи. Работы по развитию и конкретизация трехкомпонентной теории шли в двух направлениях: поиска трех веществ сетчатки и зависимости чувствительности их от длины волны; изучения чувствительности трех приемников по даиным колориметрических измерений.
Следует заметить, что чувствительность вещества| сетчатки может отличаться от чувствительности емника в целом, так как информация, полученная колбочек, довольно сложно кодируется.
5.3. Поиски светочувствительных веществ сетчатки.
Уже давно (в конце прошлого века) из сетчатки было выделено светочувствительное вещество – зрительный пурпур, или родопсин, о котором стало известно, что он выцветает под действием света, теряя светочувствительность, и постепенно восстанавливается в темноте, приобретая прежние свойства. Правда, восстановление требовало некоторых условий, которые обеспечиваются в живой сетчатке в нормальных физиологических условиях (в частности, для восстановления необходим витамин А).
Спектрофотометрические исследования показали селективность чувствительности родопсина. Спектральная кривая поглощения родопсина близка к кривой относительной спектральной световой эффективности ночного зрения. Если учесть различия условий опыта с извлеченным из сетчатки родопсином (опыт in vitro) и экспериментов с живыми испытуемыми (опыт in vivo) и сделать соответствующие поправки, обе кривые практически совпадут. Таким образом, было доказано, что родопсин – пигмент палочек, которые, как известно, не различают цветов Поиски трех пигментов, которые должны содержаться в колбочках, оказались сопряженными с большими трудностями, что можно было, впрочем, заранее предвидеть. Во-первых, колбочек в сетчатке в пятнадцать раз меныне, чем палочек, а во-вторых, их малая чувствительность к свету может объясняться и малой концентрацией колбочковых пигментов.
Выделить три колбочковых пигмента до сих пор не удалось. Уолд выделил колбочковый пигмент, названный иодопсином, из сетчаток цыплят. Их сетчатки не содержат родопсина, что связано с отсутствием ночного зрения (куриная слепота), и поэтому колбочковый пигмент у цыплят оказывается в относительно большей концентрации, чем у млекопитающих. От иодопсина до нахождения трех светочувствительных веществ колбочек было еще далеко. Хотя экстрагировать разделыю три колбочковых пигмента до сих пор не удалось, эксперименты, проведенные другими методами, показали, что в сетчатке кроме родопсина присутствуют еще три вида пигментов с разными максимумами спектральной чувствительности. Методом микрофотометрии, с помощью которого измерялась спектральная зависимость коэффициента поглощения отдельных колбочек на участках сетчатки человека и обезьяны, было показано, что зрительная система действительно характеризуется тремя видами рецепторов. Маркс, Добел и Мак Никол, а также Браун и Уолд установили наличие трех пигментов с максимумами поглощения при длинах волн 448, 558 и 567 нм.
Аналогичный результат дали и электрофизиологические исследования. Томита с сотрудпиками исследовал сетчатку карпа с помощью микроэлектродов, диаметр кончиков которых был менее 0,1 мкм. Измеряя изменения электрического потенциала при стимуляции сетчатки излучениями с различиыми длинами волн, исследователи обнаружили три типа колбочек с тремя различными спсктральными функциями. Максимальные изменения потенциала наблюдались для каждого вида колбочек при длинах волн 462, 529 и 611 нм. Таким образом, теория, трехкомпонентности цветового зрения получила экспериментальные подтверждения.
Обработка и кодирование сигналов, вырабатываемых под действием света в рецепторах, начинается уже в самой сетчатке, состоящей из несколышх слоев нервных клеток. Эти слои тоже были подвергнуты испытаниям с помощью микроэлектродов. Реакция на световой стимул при передаче сигнала в последующие слои усложняется. Появляются потенциалы как положительные, так и отрицательные, развиваются явления возбуждения и торможения (по физиологической терминологии). При перемещении электродов в слой ганглиозных клеток сетчатки регистрируются серии импульсов потенциала постоянной амплитуды, но различной частоты. Частота импульсов зависит от энергии светового стимула,
от его спектрального состава и положения электрода в слое ганглиозных клеток. Видимо, эти серии импульсов передаются в мозг, где в конечном счете и декодируются (расшифровываются), вызывая ощущенне цвета в том месте пространства, откуда исходит световой импульс.
Опыты с микроэлектродами можно считать прямым экспериментальным доказательством трехкомпонептной теории цветового зрения. Некоторую разницу в длинах волн, на которые приходятся максимумы чувствительности трех пигментов колбочек, можно объяснить погрешностью эксперимента. Да кроме того, у человека и карпа светочувствительные вещества не обязателыю должны быть совершенно одинаковыми.
5.4.Гипотеза Геринга.
Врамках трехкомпонентной тсорни делались и более сложные предположения о свойствах трех приемников глаза. Э. Геринг (1834–1918) (23) считал, что мы воспринимаем как простые шссть цветов: красный, желтый, зеленый, синий, бслый и черный, Эти цвета Геринг связал в пары: красный – зеленый (к–з), желтый – синий (ж–с) и белый – черный (б– ч). Каждой паре соответствует свое вещество, так что трехкомпонентность зрения Герингом не опровергается. Но поведепие каждого из веществ под действисм света не такое, как по теории Ломоносова – Юнга – Гельмгольца. Лучи разных частей спектра действуют различно на три спеточувствительных вещества. Длинноволиовые излучения вызывают разложеняе (дниссимиляцию) вещсств к–з и ж–с и ощущение красного или желтого соответственно. Коротковолновые излучения приводят к восстановлению (ассимиляции) этих веществ и к ощущению зелепого и синсго. Воздействие всякого видимого света разлагает б–ч – вещество, вызывая ощущепие белого. В темнотс б–ч восстанавливается.
Если отложить длины волп по оси абсцисс, а воздействие па приемники к–з и ж–с по оси ординат, мы получим две кривые, каждая из которых проходит местами выше, местами ниже оси ординат: диссимилядии соответствуют положительныс области, а ассимиляции – отрицательные. Кривая б–ч всюду положительна, так как отрицателышх яркостей не бывает. Видимо, кривая б–ч близка к кривой относительной спектральной световой эффективности
V(λ).
Гипотеза Геринга безусловно неверна для веществ сетчатки, но для объяснения явлений на более высоких уровнях, где наблюдаются возбуждения и торможения, подобная гипотеза – гипотеза оппонентности – может оказаться плодотворной. Заметим, что на Международном конгрессе «Цвет-85» (16–22 июня 1985 г. в городе Монте-Карло, пятый конгресс Международнои ассоциации по цвету – АIС) в качестве постулата принято сушествование шести «чистых» основных цветов: белого, черного, желтого, красного, голубого
изеленого.
5.5.Исследование чувствительности трех приемников с помощью колориметрических измерений.
Определенные данные о спектральной чувствительности каждого из трех приемников сетчатки оказалось возможным получить с помощью колориметрических экспериментов по сложению дветов. Существенную роль здесь сыграли опыты, пронодимые параллельно с нормальными наблюдателями (трихроматами) и лицами, у которых не работает один из приемников цвета (дихроматами). Лица, у которых не работает приемник средней части видимого спектра, называются зеленослепыми или дейтеранопами, а неспособные воспринимать длинноволновую часть спектра – краснослепыми или протанопами. Весьма редко встречаются люди, не воспринимающие коротковолновую часть спектра (тританопы).
Первые опыты с протанопом провел уже Максвелл. С дихроматами работали Ф. Питт (1935 г.), Л. Томсон и В. Райт (1953) и другие ученые. Пока мы не познакомили читателя с системами координат, в которых измеряется цвет, объяснять метод определения кривых чувствительности трех приемников глаза затруднительно. Скажем здесь только о его результатах. Наиболее точные данные получены Н. Д. Нюбергом и Е. Н. Юстовой (43, 61).
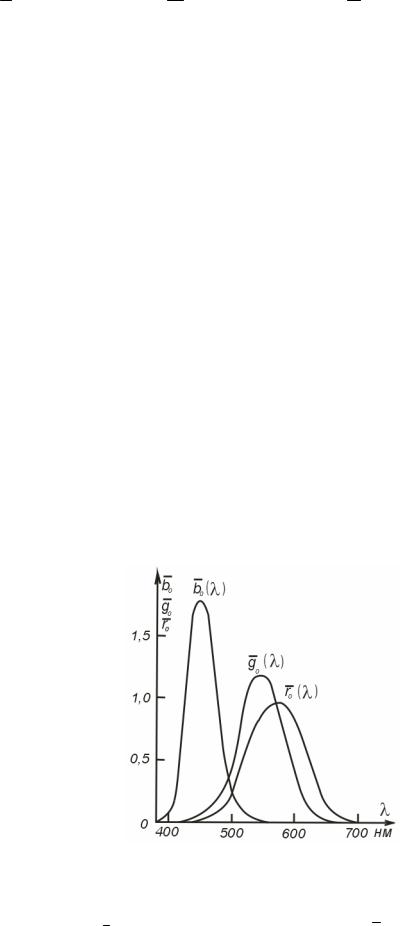
Они представлены в табл. 5.1 и на рис. 5.5, где чувствительюсть длинноволнового приемникаr0 (λ) , средневолнового g0 (λ) , коротковолнового b0 (λ) (62).
Табл. 5.1 можио принять за основу физиологической системы измерения цвета R0, R0, B0, добавив еще условие, что при равенстве координат цвета r0' = g0' = b0' получается белый равноэнергетический цвет WЕ, т. е. цвет, для которого Рλ = const для всей видимой области.
Напомним, что Pλ = |
dP |
, т. е. это спектральная плотность потока. |
|
|
||||||
dλ |
|
|
||||||||
|
|
|
|
|
|
|
|
|
||
|
Таблица 5.1 Спектральные характеристики приемников глаза среднего стандартного |
|||||||||
|
|
|
|
|
|
|
|
|
наблюдателя (1950г.) |
|
λ,нм |
|
r0 |
|
g0 |
b0 |
λ,нм |
r0 |
g0 |
b0 |
|
380 |
|
0,0000 |
|
|
0,0000 |
0,0065 |
580 |
0,9408 |
0,7672 |
0,0017 |
390 |
|
0,0001 |
|
|
0,0001 |
0,0201 |
590 |
0,8875 |
0,5621 |
0,0011 |
400 |
|
0,0002 |
|
|
0,0003 |
0,0679 |
600 |
0,8020 |
0,3734 |
0,0008 |
410 |
|
0,0004 |
|
|
0,0011 |
0,2074 |
610 |
0,6864 |
0,2258 |
0,0003 |
420 |
|
0,0013 |
|
|
0,0042 |
0,6465 |
620 |
0,5485 |
0,1273 |
0,0002 |
430 |
|
0,0036 |
|
|
0,0154 |
1,3856 |
630 |
0,3960 |
0,0663 |
0,0000 |
440 |
|
0,0081 |
|
|
0,0351 |
1,7471 |
640 |
0,2686 |
0,0330 |
|
450 |
|
0,0142 |
|
|
0,0636 |
1,7721 |
650 |
0,1672 |
0,0157 |
|
460 |
|
0,0242 |
|
|
0,1049 |
1,6692 |
660 |
0,0963 |
0,0075 |
|
470 |
|
0,0441 |
|
|
0,1554 |
1,2876 |
670 |
0,0508 |
0,0035 |
|
480 |
|
0,0817 |
|
|
0,2224 |
0,8130 |
680 |
0,0271 |
0,0017 |
|
490 |
|
0,1381 |
|
|
0,3133 |
0,4652 |
690 |
0,0131 |
0,0007 |
|
500 |
|
0,2305 |
|
|
0,4647 |
0,2720 |
700 |
0,0065 |
0,0004 |
|
510 |
|
0,3768 |
|
|
0,6980 |
0,1582 |
710 |
0,0033 |
0,0002 |
|
520 |
|
0,5566 |
|
|
0,9485 |
0,0782 |
720 |
0,0017 |
0,0001 |
|
530 |
|
0,7057 |
|
|
1,1058 |
0,0422 |
730 |
0,0008 |
0,0000 |
|
540 |
|
0,8152 |
|
|
1,1719 |
0,0203 |
740 |
0,0004 |
|
|
550 |
|
0,8903 |
|
|
1,1611 |
0,0087 |
750 |
0,0002 |
|
|
560 |
|
0,9392 |
|
|
1,0865 |
0,0039 |
760 |
0,0000 |
|
|
570 |
|
0,09569 |
|
|
0,9503 |
0,0021 |
|
|
|
|
* Яркостные коэффициенты LR0: LG0: LB0 = 1: 0,655: 0,003
Рис. 5.5. Характеристики спектральной чувствительности трех приемников глаза (по Е. Н. Юстовой).
Координаты цвета r0',g0',b0' вычисляются по формулам |
|
r0 '= ∫Pλ r0 (λ)dλ; g0 '= ∫Pλ g0 (λ)dλ; b0 '= ∫Pλb0 (λ)dλ . |
(5.2) |

Интегралы берутся в пределах всей видимой области спектра. Яркости единичных цветов характеризуются их яркостными коэффициентами LR0, LG0 и Lв0. Но абсолютные значения этих коэффициентов не определяются. Известно только их отношение друг к другу
(63)
LR0 : LG0 : Lв0, = 1: 0,655:0,003. |
(5.3) |
Однако в практике крлориметрических измерений физиологическая система |
почти. не |
применяется. |
|
5.6. О форме кривых чувcтвительности.
При изучении строения и работы органов живых существ полезно бывает задаваться вопросом не только, как (как устроен, как работает), но и вопросами, отчего и для чего. Ведь живое существо устроено рационально, и каждый его орган должен наилучшим (или почти наилучшим) образом выполнять свое назначение.
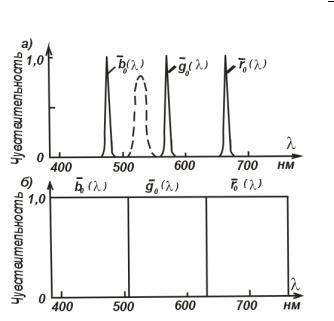
Так вот, отчего кривые спектральной чувствигельности цветовоспринимающих веществ сетчатки имеют такую форму? Для чего они пересекаются? Почему любое излучение, даже спектрально-чистое, возбуждает, как правило, два, а иногда и все три приемника? Какая форма кривых может показаться более рациональной? Что произойдет, если выбрать узкополосные, т. е. очень селективные, приемники вроде изображенных на рис. 5.6, a? Предположим в глаз попадет излучение, сосредоточенное в узкой полосе спектра вокруг λ = 530 нм (штриховая кривая на рис. 5.6). Оно не возбудит ни одного приемника, никакого цвета мы не воспримем, В лучшем случае, если излученйе попадет не в центр сетчатки, а в периферическую ее часть, где естъ палочки, мы увидим более или менее светлое пятно. А если еще глаз адаптирован к достаточно высокой яркости, когда палочки заторможены, то и вообще ничего не увидим.
Сделаем поправку. Поделим всю область видимого спектра на три равные части так, чтобы спектральные области были поровну поделены между тремя приемниками и чувствительность каждого в пределах его области была посгоянной (рис. 5.6,б) Пусть на такую систему подействует такая же полоса спектра, какая изображена на рис. 5.6, а. На нее прореагирует приемник g0 (λ) . Все как будто хорошо. Но подействуем другим излучением –
полосой спектра с центром λ = 580 нм. Реагировать будет тот же приемник g0 (λ) , и мы
увидим тот же цвет. Между тем λ = 530 нм соответствует зеленому, а λ = 580 нм – желтому, т. е. цветам, которые глаз прекрасно различает.
Изменение формы каждой кривой, скажем, на колоколообразную при сохранении каждой из них в своей области спектра не улучшит цветоразличения. Излучение λ = 580 нм вызовет большее возбуждение, чем излучение λ = 530 нм, но, поскольку возбужден будет тот же приемник g0 (λ) , глаз воспримет только большую яркость излучения, а разницы в
цветности не ощутит.
Теперь обратимся к реальной системе приемников, т. е. к рис. 5.5 и табл. 5.1. Сравним воздействие двух излучений на приемники глаза:
|
λ |
|
r0 (λ) |
g0 (λ) |
|
|
|
|
|
b0 (λ) |
|||||
1………. |
530нм |
0,7057 |
1,1058 |
0,0422 |
|||
2………. |
580нм |
0,9408 |
0,7672 |
0,0017 |
|||
В первом случае больше всего возбужден зеленый приемник. Но возбуждены и другие. Во втором случае – больше всего красный опять-таки при. возбуждении других. Сигналы от возбужденных колбочек идут в мозг и там определенным образом суммируются. В зависимости от вклада в эту сумму каждого приемника, от соотношения между степенями возбуждения каждого мы определяем цвет и тонко отличаем друг от друга даже близкие цвета. Наименьшим, еще ощутимым цветовым различиям посвящена особая глава (14). Здесь мы хотим только указать, что именно наложение кривых трех приемников обеспечивает
способность тонко различать цвета. Взглянув на рис. 5.5 и табл. 5.1, мы видим, что в крайней красной части спектра перекрытия уже нет: работает только приемник r0 (λ) .
Рис 5.6. Гипотетические кривые спектральной чувствительности приемника глаза.
И действительно, после λ = 720 нм до красного конца спектра. Излучения всех длин волн воспринимаются нами одинаково, так что, осветив одно поле фотометра светом с длиной волны 750 нм, а другое – с длиной волны, скажем, 730 нм и подобрав соответствующие яркости обоих полей, мы сможем сделать оба поля неразличимыми по цвету. Здесь наглядно обнаруживается недостаток изолированности отдельного приемника и, следователыю, преимущество перекрытия кривых чувствительности приемников. Так мы находим ответ; на вопрос, для чего сформировалась та система приемников, которая существует в действителыюсти.
5.7. Аномалии цветового зрения.
Спектральная чувствительность каждого из трех приемников глаза была получена на основе усреднения результатов измерений, проведенных с двадцатью двумя цветоаномалами. Из них было 10 протанопов (краснослепых) и 12 дейтеранопов (зеленослепых), Следует помнить, что спектральная чувствительность каждого из приемников у любого .человека отличается от усредненной кривой. В силу индивидуальных различий часто случается, что равенство по цвету двух полей колориметра, установленное одним наблюдателем, не вполне удовлетворяет другого. Однако это не значит, что цветовое зрение одного из наблюдателей ненормально, что один из них цветсн аномален. Но отличие цветового зрения от среднего можно считать нормальным только до известного предела, вне которого приходится констатировать аномалию цветового зрения.
Напомним, что человек, у которого работают всд три приемника цветового зрения, называется трихроматом. Патологическое состояние зрения, при котором один из приемников совсем не работает, называется дихромазией. О классификации дихромазии и о назваииях трех типов дихроматов уже было сказано выше. Есть люди, у которых восприятие цвета полностью отсутствует. Их называют монохроматами. Монохромат видит мир как черно-белую фотографию. По-видимому, колбочки у иего совсем не работают, сохранена только функция палочек. Многие особенности монохроматического зрения подтверждают такое предположение. Прежде всего то, что кривая относительной спектральной световой эффективности монохромата сдвинута к коротковолновой части спектра, т. е. совпадает с кривой V′(λ), см. § 3.4. Другие расстройства, обычно сопровождающие полное отсутствие
цветовых восприятий, также подтверждают, что у монохромата работают только палочки. У монохромата обычно сильно понижена острота зрения, наблюдается светобоязнь.
Кроме цветнослепых существуют люди, у которых работают все три приемпика цвета, но какой-то приемник ослаблен и представление о цветах оказывается искаженным. Такие люди пе признают цветовых равенств, установленных нормальным наблюдателем, но различение цветов по всему спектру у них сохранено. Их называют цветоаномалами: протаномалами или дейтераномалами – в зависимости от того, какой приемиик у них ослаблен. В дальнейшем всякое выходящее за пределы нормы отклонение цветового зрения мы будем называть аномалией цветового зрения, а человека с такой аномалией – цветоаномалом. Заметим, что можно в какой-то степени узнать, как видит тот или иной цвет цветоаномал, Дело в том, что встречаются, хотя и редко, люди, у которых только один глаз ненормален в смысле цветовосприятия. Сравнивая свои впечатления при наблюдении той или иной окрашенной поверхности то одним, то другим глазом, такой наблюдатель может сам оценить, какое искажение цвета создает его дефект цветовосприятия, и объяснить это искажение в понятных нам терминах.
Выдающийся английский ученый, фязик и химик Джон Дальтон (1766–1844) был протанопом. Он дал описание своему недостатку зрения, который с того времени стали называть дальтонизмом. Конечео, дальтонизмом следовало бы называть протанопию, но неспециалисты часто так называют любую аномалию цветового зрения. Аномалии цветового зрения иногда появляются в результате некоторых болезней, но чаще всего бывают врожденными, и в этих случаях они неизлечимы.
5.8. Статистические данные о цветоаномалах.
По данным Е. Б. Рабкина, среди населения Харькова ненормальности цветового зрения обнаружены у 7,4 % мужчин и у 0,8 % женщин.
В книге Д. Джадда и Г. Вышецки (23) дана таблица (табл. 5.2), составленная по результатам обследования цветового зрения 23 000 школьников.
Таблица 5.2. Распределение врожденных цветоаномалий по их типам
Дефект |
Процент имеющих указанный дефект |
|
|
мужчин |
женщин |
Протоаномалия |
1,0 |
0,002 |
Дейтероаномалия |
4,9 |
0,38 |
Протанопия |
1,0 |
0,02 |
Дейтероанопия |
1,1 |
0,01 |
Тританопия |
0,002 |
0,001 |
Полная цветовая слепота (монохромазия) |
0,003 |
0,002 |
Мы видим, что монохромазия – явление довольно редкое. Она встрсчается менее чем у одной сотой процента населения. И затем обнаруживается удивительный факт, что женщины страдают дефектами цветового зрения по меньшей мере в 50 раз реже, чем мужчины.
Объяснеиие находят в своеобразной асимметрии в наследственной передаче цветоаномалий. Для определенности рассмотрим какую–нибудь одну форму цветоаномалий, скажем дейтероформу, т. е. дейтераномалию или дейтеранопию. Женщина, имеющая в своем генетическом коде скрытые причипы дейтероформы, обычно обладает нормальным зрением. Но своему сыну она может передать соответствующий дефект. А ее дочь будет нормальным трихроматом, хотя, быть может, и унаслсдует ген дейтероформы. Но если ее отец окажется дейтераномалом илн дейтеранотом, тот же дефект проявится и у дочери. Поскольку
вероятность такого совпадения генетических кодов мала, соответствующие ненормальности у.женщин встречаются редко. Сказанное в той же мере относится и к протоаномалии.
5.9. Диагностика дефектов цветового зрения.
Выявление недостатков цветового зрения имеет серьезное практическое значение, так как нормальное различение цветов необходимо в некоторых областях профессиональной деятельности человека. Более того, допущение к некоторым профессиям цветоаномала грозит непосредственной опасностью катастроф и человеческих жертв. Это относится прежде всего к водителям транспортных средств, которые должны руководствоваться цветовыми сигналами, прсжде всего отличать красный цвет от зелеяого.
Впрошлом веке цветовая слепота водителя не раз приводила к катастрофам. Особенно много жертв повлекло крушение поезда в Лагерлунде в 1875 году: цветнослепой машинист принял красный сигнал за зеленый. Это крушение послужило толчком для организации проверок цветового зрения прежде всего у работников транспорта. Опасность увеличивается тем, что цветоаномалы и даже цветнослепые часто сами не знают о своем недостатке. Они судят о цвете по предмету, для которого он характерен, по яркостной характеристике цвета и, привыкнув к этому с детства, способны называть цвета так, как называют их окружающие. Дальтон не знал, что он протаноп, до двадцати шести лет. Все это обусловливает необходимость разработки средств проверки цветового зрения при профотборе для ряда профессий.
Проще всего цветоаномалия может быть выявлена с помощью специальных таблиц. Широко известны таблицы Ишихара. В Советском Союзе с 1936г. применяются «полихроматические таблицы для исследования цветоощущения» Е. Б. Рабкина (47). Каждая таблица заполкена пятнами разных размеров и цветов, и на их фоне человек с нормальным зренкем различает некоторые знаки, тоже образованные пятнами, несколько отличающимися по цвету. Например, на одной из таблиц видно число 96. Но протаноп различает только цифру 9, а дейтераноп – только 6.
Более точные количественные данные о цветоаномалии можно получить с помощью прибора, который называется аномалоскопом.
5.10.Аномалоскопы.
Ваномалоскопе испытуемый видит рядом два поля, которые он должен уравнять по цвету. Обычно цвет одного из полей остается неизменным, а спектральный состав света, падающего на другое поле, можно менять.
Наиболее известен аномалоскоп Нагеля, построенный в 1907 году на основе исследований Релея, опубликованных еще в 1882 году. В нем одно поле освещается желтой линией натрия 589 нм (точнее дублет – 589,0 и 589,6 нм), а другое – смесью красной линии лития 671 нм и зеленой таллия 536 нм, Количество красного и зеленого излучений можно менять, изменяя ширину щелей, пропускающих освещающий поле свет, Яркость желтого поля тоже можно регулировать.
Испытуемому предоставляется путем вращения барабанов установить равенство обоих полей по цвету. Люди с нормальным цветовым зрением устанавливают такое равенство при вполне определенном отношении интенсивностей зеленого и красного. Точнее говоря, различия в их установках существуют, но они невелики и лежат в некоторых пределах, которые и характеризуют норму. Цветоаномал принимает установку нормального испытуемого, но готов принять и другие, сильно отличающиеся установки, требуя для уравнения по цвету только изменения яркости желтого поля. Испытуемый, у которого один из приемников совсем не работает (протаноп или дейтераноп), принимает за желтый цвет любое сочетание красного и зеленого, находя для каждого сочетания определенную яркость желтого. Но отношение красного и зеленого у протанопа и дейтеранопа различно, что и дает возможность установить характер цветоаномалии. Если все три приемника работают, но один из них ослаблен, испытуемый способен уравнять с желтым не все соотношения
красного и зеленого, допуская отклонения от нормы только в известных пределах. Чем шире эти пределы, тем сильнее цветовая аномалия испытуемого.
Аномалоскоп, основанный на совершенно новом принципе, создан Г. Н. Раутианом. В основу своей разработки Раутиан положил представление о трех приемниках цвета, спектральная чувствительность которых была определена Юстовой, Для испытания каждого из приемников была выбрана своя система фильтров, свой этап испытания цветового зрения.
В аномалоскопе Раутиана свет от одного источника освещает два омежных поля, которые испытуемый должен уравнивать по цвету. В пучки, освещающие правое и левое поля, вводятся цветные фильтры. Без них освещенности обоих полей были бы равны.
Обозначим спектральную плотность энергетической освещенности Еλ, считая, что Eλ = dEdλ .
Установка фильтра со спектральным пропусканием τ(λ) изменит эту освещенность. По аналогии с формулой (5.2) мы можем написать
r0 = ∫Eλτ(λ) |
r0 (λ)dλ; g0 = ∫Eλτ(λ)g0 (λ)dλ; |
|
|
|
|
(5.4) |
b0 = ∫Eλτ(λ)b0 (λ)dλ . |
||||||
Здесь r’0, g’0, b’0 – составляющие цвета поверхности.
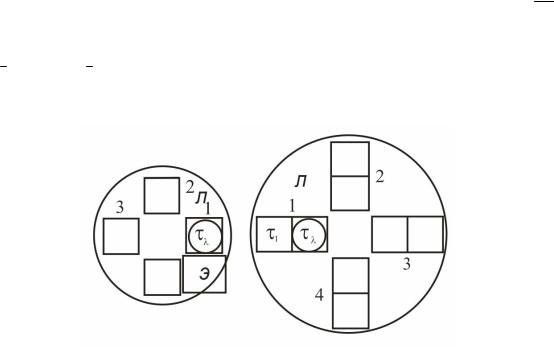
Рис. 5.7 Схема установки светофильтров в анамалоскопе Раутина.
На рис. 5.7 схематически изображена система установкп фильтров в аномалоскопе Раутиана. Все фильтры расположены на дисках Л и П. Круги внутри двух квадратов изображают сечения пучков, освещающих правую и левую части поля, которое наблюдает исследуемый. Диск Я может смещаться вдоль горизонтали вправо и влево. В начале измерения в оба пучка света введены одинаковые фильтры τλ и оба поля сравнения имеют один и тот же цвет. Затем экспериментатор начинает сдвигать диск П вправо, вводя в правый пучок все болйшую частъ фильтра τ1, отличающегося от фильтра τλ. Цвет правого поля меняется, но аномал может этого не замечать. Есля он замечает только изменение яркости, экспериментатор компенсирует ее изменение, выдвигая в левый пучок заслонку Э. Чем больше аномалия, тем дольше испытуемый не замечает разницы в цвете.
Особенность аномалоскопа Раутиаяа заключается в прдборе фильтров τλ и τ1. При введении фильтра τ1 меняется только одна составляющая цвета, например, r′0. Чем больше составляющая r′0 в правом пучке, тем краснее правое поле, видимое испытуемым. Следовательно, его испытывают на протаномалию. Кончив этот первый этап испытания, поворачивают диски П и Л, вводя в работу другое сочетание фильтров, чтобы на втором этапе провести испытание, скажем, на дейтераномалию.
Вернемся к первому этапу испытания. Человек с нормальным зрением заметит изменение цвета при введении в правый пучок некоторой части nн фильтра τ1. Аномал позволит ввести большую часть фильтра τ1, скажем, na. Конечно, и человек с нормальным зрением допустит введение некоторой части (na) фильтра τ1 до того как заметит изменение цвета. Протаномал допустит большее введение фильтра Степень цветоаномалии можно характеризовать отношением N = nа/nн.