

Семинар на 15.05.2024
Основные этапы гистогенеза поперечно-полосатой мышечной ткани
Миогистогенез или регенерация скелетной мышечной ткани
В своём развитие скелетная мышечная ткань проходит несколько этапов:
На этапе закладки мышцы, из миотомов самитов выселяются клетки миобласты веретеновидной формы и мигрируют в места бушующих мышц. Первая стадия миобласт.
На второй стадии миобласты делятся и дифференцируются в двух направлениях. Первое направление развития миосателитоциты (это клетки, которые будут находится между двумя листками сарколеммы) и второе направление развития динамическое (если миосателитоциты в таком состоянии заснут и будут находится так практически в течении всей жизни, то второе направление из миобласты будет развиваться, оно даёт толчей дальнейшему развитию). Из миобластов дальше при втором направлении развиваются миосимпласты (миобласты сливаются вместе, образуя миосимпаласт).
Миосимпласты начинают дифференцироваться, в них образуется органоиды (общие, специальные) и постепенно из миосимпласта образуются мышечные трубочки – миотубы. В миотубах ядра находятся в центре в виде цепочки, миофибриллы лежат по периферии.
Миотубы дифференцируются, миофибрилл становится больше, миофибриллы выгоняют, вытесняют ядра из центра, они уходят на периферию и формируется последняя стадия, стадия зрелого мышечного волокна.
Строение мышечного волокна как структурно-функциональной единицы скелетной мышечной ткани
С
 труктурно
функциональной единицей скелетной
мышечной ткани является мышечное
волокно. При световой микроскопии
мышечное волокно представляет собой
тяж, несколько закруглённый или
заострённый на концах.
труктурно
функциональной единицей скелетной
мышечной ткани является мышечное
волокно. При световой микроскопии
мышечное волокно представляет собой
тяж, несколько закруглённый или
заострённый на концах.Длинна этого тяжа: нижняя граница от 1-40мл верхняя граница до 150мл. Длина определяется длиной мышцы, из которой волокно было извлечено. Так, например, самые длинные мышечные волокна находятся в мышцах свободных нижних конечностей, в то же время в противовес более короткие мышечные волокна находятся в мышцах кисти, а так же в среднем ухе (мышца, натягивающая барабанную перепонку и стременная мышца). Диаметр мышечного волокна от 9-150мкм.
М
 ышечное
волокно с поверхности покрыто оболочкой,
которая носит название сарколемма. Под
сарколеммой находится аналог цитоплазмы
– саркоплазма.
ышечное
волокно с поверхности покрыто оболочкой,
которая носит название сарколемма. Под
сарколеммой находится аналог цитоплазмы
– саркоплазма.
По периферии саркоплазмы, сразу под сарколеммой так же на светооптическом уровне, при световой микроскопии, можно увидеть многочисленные ядра, а значит мышечное волокно представляет собой НЕ клеточную форму организации живой материи, а именно мышечный симпласт, миосимпласт. Так же при световой микроскопии, но на чуть большем увеличении мы сможем увидеть миофибриллярную поперечную исчерченность.
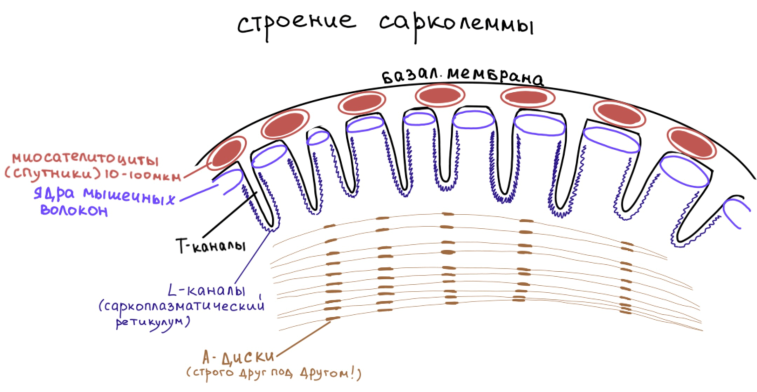
При электронно-микроскопическом исследовании было установлено, что оболочка мышечного волокна имеет двухслойное строение. Наружный ее листок построен по типу базальной мембраны, внутренний листок построен по типу клеточной мембраны. При этом внутренний листок формирует очень глубокие инвагинации, которые ориентированы поперёк длинной оси мышечного волокна. Эти инвагинации получают название Т-каналов. Между двумя листками сарколеммы располагаются малодифференцированные клетки, – миосателитоциты или мышечные спутники. Они находятся здесь ещё с эмбрионального периода и именно эти клетки определяют возможность регенерации мышечных волокон по клеточному типу.
Миосателитоциты представляют собой малодифференцированные клетки, это стволовые клетки овальной формы, размером 10-100мкм, с высоким ядерно-цитоплазматическим соотношением, цитоплазмы мало, органоиды развиты слабо. Соотношение между миосателитоцитами и ядрами мышечного волокна (они находятся под сарколеммой) составляет 1:10, то есть на один миосателитоцит приходится 10 ядер мышечного волокна.
В мышечных волокнах можно выявить как общие, так и специальные органоиды. К общим относят митохондрии (развиты хорошо), гладкую ЭПС (развита хорошо), она, подходя к Т-каналам, начинает закручиваться вокруг последних и образует L-каналы (продольные), предназначение L-каналов сводится к депонированию ионов кальция (здесь накапливаются ионы кальция, необходимые для сокращения). Другое название L-каналов – саркоплазматический ретикулум.
Гранулярная ЭПС и комплекс Гольджи редуцированы и их остатки находятся около полюсов ядер. Данные органоиды были хорошо развиты на более ранних стадиях формирования мышечных волокон.
С
 пециальные
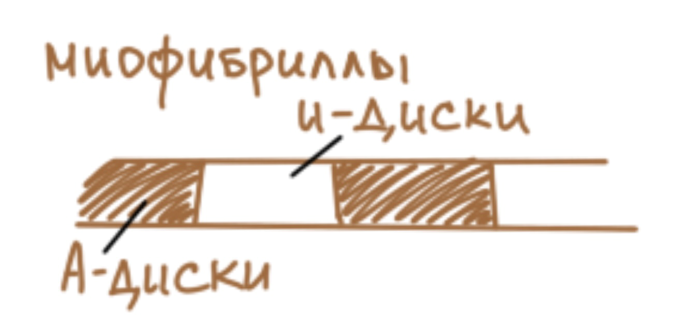
органоиды. Это миофибриллы или
сократительные нити. Миофибриллы
занимают всю центральную часть мышечного
волокна, вытесняя ядра на периферию. В
составе каждой миофибриллы можно
выделить темные участки и светлые
участки. Темные участки получают
название А-дисков от термина анизотропные
диски. Это диски, обладающие двойным
лучепреломлением в поляризованном
свете. Светлые диски получают название
И-диски от термина изотропные диски,
они обладают одинарным лучепреломлением
в поляризованном свете. Все А-диски
находятся строго друг под другом, точно
так же, как и все И-диски, в результате
чего, на уровне мышечного волокна даже
при световой микроскопии мы видим
миофибриллярную исчерченность.
пециальные
органоиды. Это миофибриллы или
сократительные нити. Миофибриллы
занимают всю центральную часть мышечного
волокна, вытесняя ядра на периферию. В
составе каждой миофибриллы можно
выделить темные участки и светлые
участки. Темные участки получают
название А-дисков от термина анизотропные
диски. Это диски, обладающие двойным
лучепреломлением в поляризованном
свете. Светлые диски получают название
И-диски от термина изотропные диски,
они обладают одинарным лучепреломлением
в поляризованном свете. Все А-диски
находятся строго друг под другом, точно
так же, как и все И-диски, в результате
чего, на уровне мышечного волокна даже
при световой микроскопии мы видим
миофибриллярную исчерченность.
Строение миофибриллярного аппарата скелетной мышечной ткани
Гистофизиология мышечного сокращения
Сокращение мышечного волокна начинается с того, что на мышечное волокно поступает нервный импульс, который приходит по аксону двигательного нейрона из передних рогов спинного мозга, при этом на мышце образуется нервно-мышечный синапс или двигательное нервное окончание.
Поступив на мышечное волокно нервный импульс начинает одновременно проходить сначала по всем Т-каналам, после чего переходит на L-каналы, что приводит к высвобождению из L-каналов ионов кальция.
Когда концентрация ионов кальция в саркоплазме достигает пороговой величины, начинаются изменения пространственного строения белков, входящих в состав актиновой и миозиновой протофибрилл. При этом на актиновой протофибрилле меняется конформация тропомиозина, что приводит к смещению с тропомиозина глобул тропонина. В результате чего открываются активные центры на актиновой протофибрилле.
Под действием ионов кальция происходят конформационные изменения и с миозиновой протофибриллой, а именно активируется белок миозинатфаза и начинается гидролиз/распад молекул АТФ. В результате чего миозиновая головка имеет возможность присоединиться к активному центру.
Происходит адгезия активной головки к активному центру на активной протофибрилле (т. е. между ними мости образуются связывающие, они, грубо говоря, приклеиваются друг к другу).
Параллельно с этим, высвободившаяся энергия, меняет пространственное строение шарнирного участка, и миозиновая головка начинает совершать гребковые движения и начинает сдвигать актиновую нить вдоль миозиновой до тех пор, пока продолжает поступать нервный импульс и до тех пор, пока в саркоплазме присутствует кальций.
На уровне саркомера при этом наблюдается смещение актиновых нитей к центру саркомера и укорочение И-дисков (А-диски не меняются, потому что скользят актиновые вдоль миозиновых) и укорочение длинны всего саркомера. Тянущее усилие через Z-линии передаётся на сарколемму, что приводит к сокращению мышечного волокна целиком.
Для расслабления мышечного волокна необходимо прекращение поступления нервных импульсов, а кальций должен вернуться обратно в саркоплазматический ретикулум.
Для того чтобы вернуть обратно кальций необходима энергия АТФ. В L-каналах кальция и так много, а раз он будет возвращаться туда, где его и так много – это против градиента концентрации транспорт, активный транспорт.
Как только концентрация кальция в саркоплазме уменьшится, миозиновые головки отсоединятся от активных центров, изменится конфирмация белка тропомиозина и активные центры вновь закроются тропонином, мышечное волокно расслабится.
Строение мышцы как органа
П
 редставляет
собой комплекс мышечных волокон
различного типа, находящихся в тесной
морфофункциональной связи с элементами
соединительной ткани.
редставляет
собой комплекс мышечных волокон
различного типа, находящихся в тесной
морфофункциональной связи с элементами
соединительной ткани.
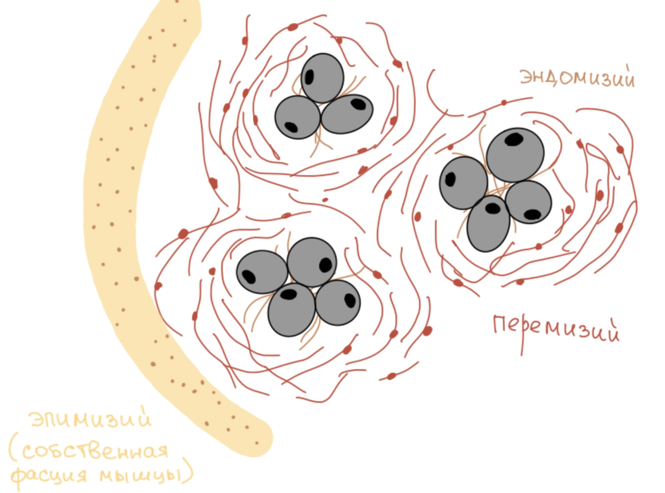
Мышечные волокна, группируясь, образуют пучки различных порядков. Между отдельными мышечными волокнами проходят очень тонкие и нежные прослойки из рыхлой соединительной ткани (РВСТ), которые получают название эндомизий.
В составе эндомизия проходят тончайшие сосуды, а так же нервные окончания, таким образом, эндомизий выполняет трофическую функцию.
Между пучками мышечных волокон, а так же вокруг них проходят более выраженные прослойки рыхлой волокнистой соединительной ткани, которые несут в себе сосуды уже более крупного калибра, а так же нервные стволики. Данные прослойки, разделяющие пучки и окружающие их, носят название перемизий. Функция тоже трофическая.
В то же время, в совокупность всех мышечных пучков с их соединительной тканью, поверхность окружается плотной соединительнотканой оболочкой, которая называется эпимизий.
Эпимизий для мышцы представляет ее вспомогательный аппарат или собственную фасцию мышцы.
Т
 ипы
мышечных волокон скелетной мышечной
ткани
ипы
мышечных волокон скелетной мышечной
ткани
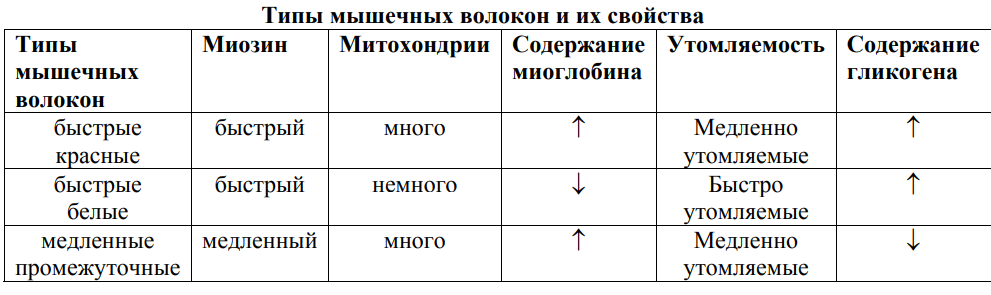
Красные мышечные волокна |
Белые мышечные волокна |
Тонкие |
Толстые, т. е. у белых диаметр больше |
В красных волокнах много белка миоглобина (аналог гемоглобина, он запасает кислород, когда мышцы расслаблены) |
В белых волокнах миоглобина мало |
Много митохондрий |
Мало митохондрий |
Включения. Преобладают липиды |
Включения. Преобладает гликоген |
Ферменты. Окислительновосстановительные, т. е. кислородозависимые |
Ферменты. Гликолитический, анаэробные |
По функциональным особенностям красные волокна являются тоническими (долгое время не утомляются) |
Белые волокна тетонические (характерны для коротких вспышек мышечной активности, т.е. быстро утомляемые) |

Физиологическая и репаративная регенерация скелетной мышечной ткани в свете теории дифферонного строения ткани
Физиологическая регенерацияпроявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличение числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что в конечном счете проявляется увеличением функциональной способности мышечного волокна.
Репаративная регенерацияразвивается после повреждения мышечных волокон. При этом способ регенерации зависит от величины дефекта. При значительных повреждениях на протяжении мышечного волокна миосателлиты в области повреждения и в прилежащих участках растормаживаются, усиленно пролиферируют, а затем мигрируют в область дефекта мышечного волокна, где выстраиваются в цепочки, формируя миотрубку. Последующая дифференцировка миотрубки приводит к восполнению дефекта и восстановлению целостности мышечного волокна. В условиях небольшого дефекта мышечного волокна на его концах, за счет регенерации внутриклеточных органелл, образуютсямышечные почки, которые растут навстречу друг другу, а затем сливаются, приводя к закрытию дефекта. Однако, репаративная регенерация и восстановление целостности мышечных волокон могут осуществляться при определенных условиях: во-первых, при сохраненной двигательной иннервации мышечных волокон, во-вторых, если в область повреждения не попадают элементы соединительной ткани(фибробласты).Иначе на месте дефекта мышечного волокна развивается соединительно-тканный рубец.
Г
 ладкая
мышечная ткань: источники развития,
строение миоцитов, регенерация,
иннервация
ладкая
мышечная ткань: источники развития,
строение миоцитов, регенерация,
иннервация
Относится по гистогенетической классификации относится к мезенхимальной.
По строению к неисчерченной, по функциональным особенностям к непроизвольной тонической.
Функция: обеспечение движения внутри организма.
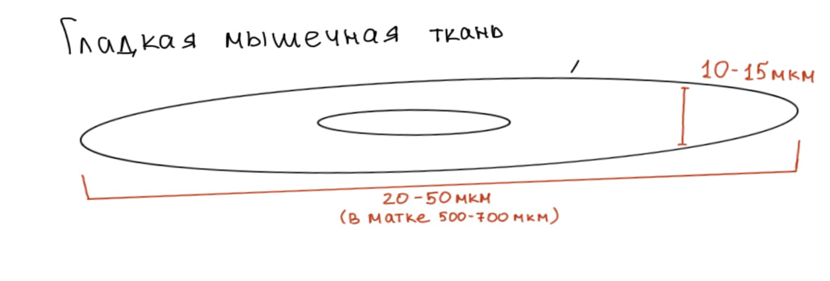
Структурно функциональной единицей гладкой мышечной ткани является клетка веретеновидной формы, называемая гладким миоцитом.
Размеры: длинна гладкого миоцита имеет размер чаще всего 20-50мкм, диаметр 5-8 – 10-15 мкм. В то же время надо понимать, что в таком органе как матка при беременности длинная гладкого миоцита может достигать 500-800 и даже 1000 мкм.
Г
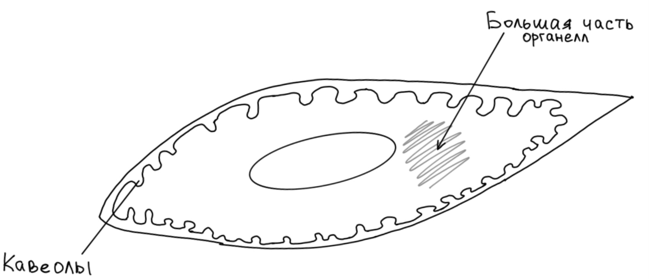 ладкий
миоцит с поверхности покрыт двухслойной
клеточной оболочкой, наружный листок
которой построен по типу базальной
мембраны, а внутренний по типу плазмалеммы.
Внутренний листок формирует неглубокие
инвагинации, которые в данном типе
клеток носят название кавеолы. В кавеолах
происходит депонирование ионов кальция.
ладкий
миоцит с поверхности покрыт двухслойной
клеточной оболочкой, наружный листок
которой построен по типу базальной
мембраны, а внутренний по типу плазмалеммы.
Внутренний листок формирует неглубокие
инвагинации, которые в данном типе
клеток носят название кавеолы. В кавеолах
происходит депонирование ионов кальция.
В центре гладкого миоцита находится палочковидное или сигарообразное ядро. Цитоплазма гладкого миоцита содержат как общие, так и специальные органоиды. Общие: митохондрии, гладкая и гранулярная ЭПС, комплекс Гольджи (среди всех мышечных элементах в гладких миоцитах синтетический аппарат развит лучше всего, это их особенность. Именно по этому гладкие миоциты могут проявлять функции фибробластов и вырабатывать компоненты межклеточного матрикса, т. е. как фибробласты вырабатывать коллаген, эластин, ГАГ, протоуглеканы и гликопротеины, секретировать и образовывать, по сути, межклеточное в
 ещество
соединительной ткани). Вышеперечисленные
органоиды находятся в цитоплазме у
полюсов ядра, в зоне эндоплазмы.
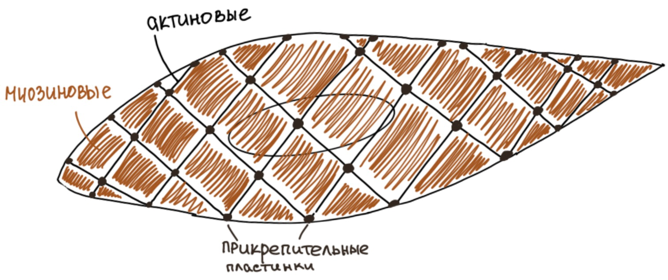
Специальные органоиды: актиновые и
миозиновые протофибриллы. Миофибрилл
нет!!
ещество
соединительной ткани). Вышеперечисленные
органоиды находятся в цитоплазме у
полюсов ядра, в зоне эндоплазмы.
Специальные органоиды: актиновые и
миозиновые протофибриллы. Миофибрилл
нет!!Актиновые протофибриллы тонкие и располагаются преимущественно вдоль длинной оси гладкого миоцита, образуя сеть. В тех участках, где актиновые протофибриллы пересекаются друг с другом и прилежат к оболочке клетки, образуются платные тельца или прикрепительные пластинки. Они являются аналогами телофрагм. Между актиновые протофибриллами располагаются миозиновые протофибриллы, но они деполимеризованы (разобраны). Именно по этому они не могут установить контакт с актиновые и сформировать миофибриллы, потому что они разобраны.
Регенерация. Гладкая мышечная ткань относится к растущим или развивающим тканям и регенерирует преимущественно по внутриклеточному типу. В то же время при определенной стимуляции, например, гармонями существует регенерации и по клеточному типу.
Иннервация. Иннервация гладкой мышечной ткани осуществляется вегетативной нервной системой – симпатическими и парасимпатическими нервными волокнами. Импульс приходит на центрально расположенную клетку мышечного пучка, все остальные клетки стимул к сокращению получают за счёт щелевидных контактов или нексусов. Сокращение мышечного пучка начинается с отшнуровывания кавеол и высвобождения из них ионов кальция. Под действием ионов кальция полимеризуется миозин, который вступает в соприкосновение с актином и только сейчас на короткий промежуток времени, только в момент сокращения могут формироваться подобия миофибрилл. При этом актиновые нити скользят вдоль миозиновых, тянущее усилие через прикрепительные пластинки передается на оболочку и гладкий миоцит начинает сокращаться (скручиваться). Ядро, расположенное между актиновых протофибриллами так же, пассивно сминается.
С
 ердечная
поперечно-полосатая мышечная ткань:
источник развития, особенности строения
и регенерации
ердечная
поперечно-полосатая мышечная ткань:
источник развития, особенности строения
и регенерации
По происхождению относится к целомическим тканям, по строению к исчерченным, по функциональным особенностям к автономным.
Иннервируется вегетативной нервной системой. В ходе эмбриогенеза развивается дивергентно (в 2х направлениях). Первое направление приводит к формированию типичной (или сократительной) сердечной мышечной ткани, второе направление приводит к формированию атипичной (проводящей) сердечной мышечной ткани.
Структурной единицей сердечной мышечной ткани (типичной или сократительной) является клетка, называемая кардиомиоцитом.
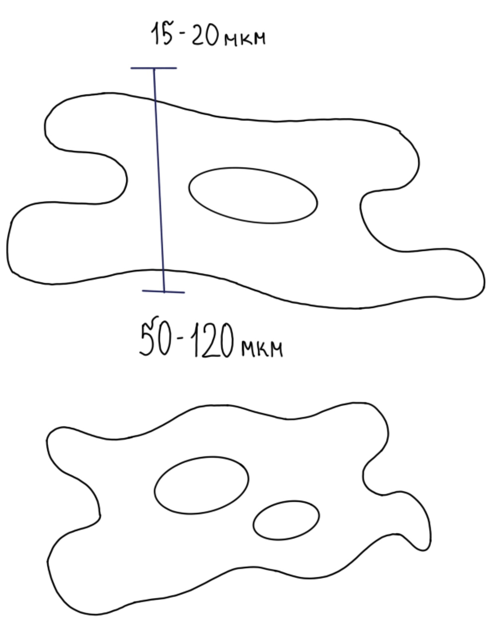
Кардиомиоцит имеет призматическую, часто отросчатую форму, причём установлено, что отростчатых кардиомиоцитов содержится больше в предсердии и несколько меньше в желудочках.
Размеры кардиомиоцитов: длина находится в пределах от 50 до 120 мкм, диаметр – 15–20 мкм.
Кардиомиоцит покрыт 2х-слойной оболочкой, где наружный листок построен по типу базальной мембраны, внутренний листок построен по типу плазмалеммы. При этом внутренний листок формирует неглубокие инвагинации, ориентированные поперёк линии, как кардиомиоцит. Данные инвагинации называются Т-каналами.
Кардиомиоциты имеют расположенное в центре одно или два ядра, а также органоиды общего и специального назначения. Среди общих органоидов в кардиомиоцитах выделяют хорошо развитые митохондрии (очень много), гладкую и гранулярную ЭПС (гладкая ЭПС превалирует), комплекс Гольджи.
Гладкая ЭПС идёт вдоль длинной оси кардиомиоцитов, закручивается вокруг Т-каналов и образует соответственно L-каналы. Их предназначение такое же: депонирование ионов кальция.
С
 пециальные
органоиды представлены
сократительными нитями – миофибриллами,
которые располагаются по периферии
кардиомиоцита и имеют такое же строение,
как и скелетные мышечные волокна
(характеризуются чередованием темных
и светлых участков – А- и I-дисков).
Фибриллы в кардиомиоцитах более
тонкие и нежные, поэтому исчерченность
на сердечных мышечных волокнах
прослеживается чуть слабее.
пециальные
органоиды представлены
сократительными нитями – миофибриллами,
которые располагаются по периферии
кардиомиоцита и имеют такое же строение,
как и скелетные мышечные волокна
(характеризуются чередованием темных
и светлых участков – А- и I-дисков).
Фибриллы в кардиомиоцитах более
тонкие и нежные, поэтому исчерченность
на сердечных мышечных волокнах
прослеживается чуть слабее. Кардиомиоциты соединяются друг с другом торцевыми и латеральными поверхностями, в результате чего из структурных единиц образуются структурно-функциональные единицы, которые получили название сердечных мышечных волокон.
Поскольку один и тот же кардиомиоцит своими отростками участвует в образовании сразу нескольких волокон, на препаратах не удаётся обнаружить, где начинается и где заканчивается сердечное мышечное волокно (создаётся картина трёхмерного соклетия). В тех участках, где кардиомиоциты соединяются друг с другом, образуются специфические структуры, имеющиеся только в сердечной мышечной ткани – вставочные диски или вставочные пластинки.
При электронной микроскопии было обнаружено, что в составе вставочного диска находится несколько типов межклеточных контактов, в том числе: Десмосома, Щелевидный контакт, Контакт по типу замка.
На светооптическом уровне комплекс этих контактов выявляется как косые полоски, идущие поперёк сердечных мышечных волокон. Между сердечными мышечными волокнами находятся прослойки из РВСТ, содержащие сосуды, нервы, нервные окончания, лимфоидные фолликулы (лимфоидные пузырьки).
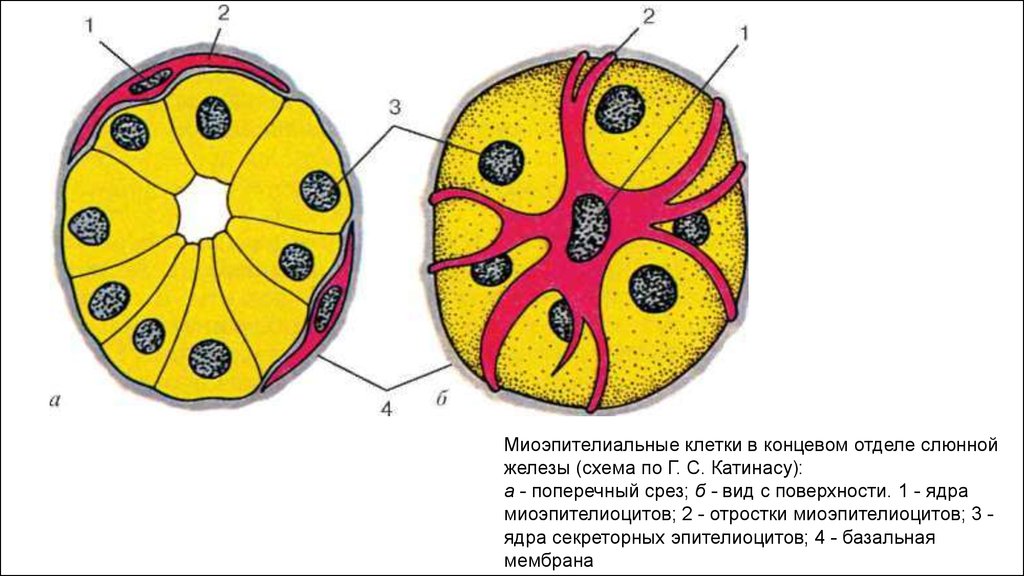
Миоэпителиальные клетки: источник развития, расположение, строение, значение
Миоэпителиальные клетки — это специализированные мышечные клетки, встречающиеся в различных тканях и органах тела, особенно в структурах, содержащих железистые или секреторные элементы.
Миоэпителиальные клетки развиваются из эпидермального зачатка.
Миоэпителиальные клетки встречаются: слюнных железах (способствуют выделению слюны в полость рта), молочных железах (окружают альвеолы и протоки, способствуя изгнанию молока во время грудного вскармливания), молочных протоках (помогают направлять молоко к соску), потовых железах (помогают выталкивать пот на поверхность кожи, помогая регулировать температуру тела), слезных железах (способствуя секреции слез), предстательной железе (помогая высвобождению простатической жидкости), бронхиолах (в стенках бронхиол дыхательной системы, помогая выводить слизь и другие выделения из дыхательных путей), экзокринной части поджелудочной железы(в протоках поджелудочной железы и способствуют перемещению пищеварительных ферментов в тонкую кишку)
Миоэпителиальные клетки уникальны тем, что сочетают в себе черты двух разных типов клеток: мышечных клеток, называемых миоцитами, и эпителиальные клетки. Такое сочетание свойств дает миоэпителиальным клеткам способность сокращаться и проявлять механическую силу, подобно мышечным клеткам, и соединяться, как эпителиальные клетки. Они отвечают за перемещение выделений из железистых клеток в протоки, ведущие к различным отверстиям или полостям тела.
Мионевральные элементы: источник развития, расположение, строение, значение