

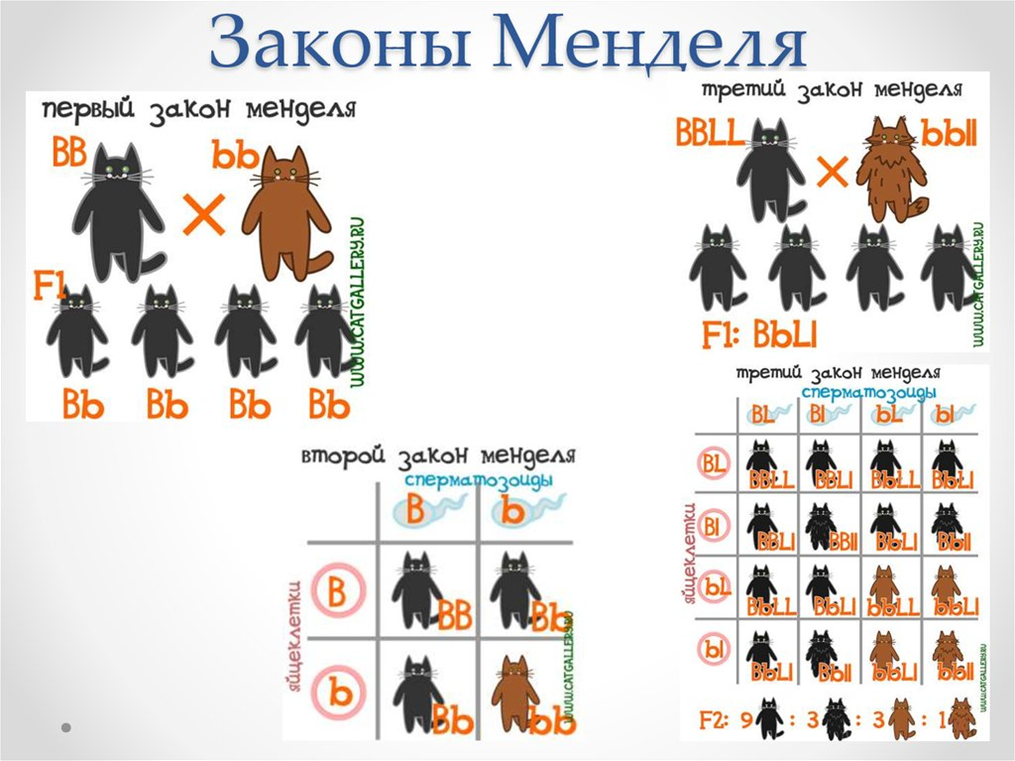
2. Закон расщепления признаков
При скрещивании двух гетерозиготных потомков первого поколения между собой во втором поколении наблюдается расщепление в определенном числовом отношении: по фенотипу 3:1, по генотипу 1:2:1.
Чистая линия - это совокупность организмов, которые на протяжении многих поколений, при скрещивании друг с другом, проявляют одинаковые признаки (не дают расщепления).
Закон чистоты гамет — в каждую гамету попадает только один аллель из пары аллелей данного гена родительской особи.
Анализирующее скрещивание – скрещивание с рецессивной гомозиготой, позволяет определить генотипы особей, не различающихся по фенотипу.
3. Закон независимого наследования признаков
При скрещивании двух гомозиготных особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании). Расщепление по фенотипу: 9:3:3:1
*Если тяжело, то тут есть ещё объяснение: https://zaochnik.com/spravochnik/biologija/genetika/zakony-g-mendelja/
3. Цитологические основы законов Менделя. Мейоз.
Цитологические основы законов Менделя включают следующие положения:
1. Соматические клетки имеют, как правило, диплоидный (двойной) набор хромосом, что означает, что аллельные гены — парные.
2. Во время мейоза, когда образуются половые клетки (гаметы), в каждую из них попадает лишь одна из пары хромосом — один аллельный ген из каждой пары.
3. Гомозиготная особь может дать только один сорт гамет — с доминантным или рецессивным признаком.
4. Гетерозигота дает два сорта гамет в равных количествах — 50 % гамет с доминантным признаком и 50 % — с рецессивным
4. Условия выполнения законов Менделя.
для 1 закона: моногенность признака. Это признак, развитие которого определяется действием одного гена. Если за фенотипический признак отвечает более одного гена (как абсолютное большинство), он всегда имеет более сложный характер наследования.
Для второго: расщепление 3:1 по фенотипу и 1:2:1 по генотипу выполняется приближенно и лишь при следующих условиях:
Большое число скрещиваний (потомков).
Равная жизнеспособность гамет, содержащих аллели А и а.
Равная вероятность слияния гамет с разными аллелями.
Одинаково жизнеспособны зиготы (зародыши) с разными генотипами.
для третьего: Закон независимого наследования выполняется при условии не сцепленного наследования. Несцепленные признаки — это признаки, гены которых находятся в разных хромосомах и наследуются независимо. Примеры несцепленных признаков: группа крови (I) и резус-фактор (R).
5. Взаимодействие генов: эпистаз, плейотропия, полимерия, комплементарность и другие примеры отклонений.
Неаллельные взаимодействия генов обусловливают отклонения:
Комплемента́рное (дополнительное) действие генов — тип взаимодействия неаллельных генов, при котором они дополняют друг друга, а признак формируется лишь при одновременном действии двух доминантных генов, которые по отдельности не приводят к появлению признака. Пример: окраска цветков у душистого горошка (Фиолетовая окраска появляется только при наличии в генотипе растения двух доминантных генов (A) и (B))
Эпистаз – доминантный ген подавляет действие неаллельного ему другого доминантного гена. У тыквы доминантный аллель A обусловливает белую окраску тыквин вне зависимости от того, какими аллелями представлен ген B.
Рецессивный эпистаз — это тип взаимодействия, при котором рецессивный аллель одного гена в гомозиготном состоянии подавляет действие доминантного аллеля другого гена. Домовые мыши с генотипом А_bb и ааbb имеют одинаковый фенотип — все белые.
Полимерия – явление, при котором развитие признаков контролируется несколькими неаллельными генами, располагающимися в разных хромосомах. Цвет кожи у человека определяют четыре пары неаллельных генов, ответственных за выработку пигмента меланина. Чем больше имеется доминантных генов, тем больше в коже меланина, и тем темнее её окраска.
Плейотропный эффект – это влияние одного гена на развитие и проявление в фенотипе нескольких признаков. Ген, обусловливающий цвет лепестков, одновременно влияет на окраску других частей цветкового растения;
6. Понятия гена. Хромосомная теории наследственности
Ген – транскрибируемый фрагмент, кодирующий белок или функциональную РНК. Имеет промотор с сайтом посадки РНК-полимеразы и терминатор транскрипции
Основные положения хромосомной теории (1911 г):
Гены локализованы в хромосомах, расположены линейно в фиксированной последовательности
Каждый ген имеет определенное место – локус на хромосоме, в идентичных локусах гомологичных хромосом находятся аллельные гены.
Гены относительно стабильны
Гены могут изменяться (мутировать)
Гены одной хромосомы наследуются совместно, образуя группу сцепления. Число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида.
Сцепление генов может быть нарушено в процессе кроссинговера – обмена участками сестринских хроматид, что приводит к образованию рекомбинантных хромосом с новыми комбинациями генов. Больше расстояние между генами – чаще кроссинговер.
7. Строение и виды хромосом. Упаковка ДНК в хромосомах.
Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками H2А, H2B, H3 и H4. Молекула ДНК «обёртывается» вокруг ядра из восьми гистоновых белков (или октамер). ДНК сложена ~ всемеро. Основная функция нуклеосомы — освобождение места в ограниченном пространстве ядра для ДНК.
Н1 гистон обеспечивает связь между нуклеосомами, образуя подобие винта (соленоида) из нуклеосом с d~300. Сам соленоид также закручивается винтом и далее - образуется иерархическая система спиралей. Гетерохроматиновые участки хромосом (конденсированные) более компактны, чем эухроматиновые.
ГЕТЕРОХРОМАТИН - участки хромосом, характеризующиеся плотной упаковкой ДНК и остающиеся в таком же состоянии после деления клеток в ходе митоза
ЭУХРОМАТИ́Н - участки хромосом, сохраняющие деспирализованное состояние в покоящемся ядре (в интерфазе) и спирализующиеся при делении клеток (в профазе); содержат большинство генов и потенциально способны к транскрипции.
Кариотип – совокупность всех хромосом, специфичность морфологии и числа хромосом. Каждый вид имеет характерный для него набор
Различают четыре типа строения хромосом:
телоцентрические (палочковидные хромосомы с центромерой на проксимальном конце)
акроцентрические (палочковидные хромосомы с очень малым вторым плечом);
субметацентрические (с плечами неравной длины, типа L);
метацентрические (V-образные хромосомы, обладающие плечами равной длины).

Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода.
Политенные хромосомы - это большие хромосомы, которые имеют тысячи нитей ДНК. Они обеспечивают высокий уровень функции некоторых тканей, таких как слюнные железы насекомых. Образуются в результате множественных циклов репликации, при котором не происходит разделения хромосом. Состоят из дисков (конденсированный хроматин), междисковых участков (более рыхлая упаковка фибрилл хроматина) и пуфов.
Пуфы политенных хромосом - это утолщения на политенных хромосомах. Они являются результатом деспирализации функционально активных участков хромосом с образованием открытых петель ДНК. Отражает повышенную транскрипционную активности.
Хромосомы типа ламповых щеток — это специальная форма хромосом, которую они приобретают в растущих ооцитах (женских половых клетках) большинства животных, за исключением млекопитающих. Боковые петли, отходящие от основной оси хромосомы. Являются сайтами активной транскрипции, наблюдаемый на стадии формирования желтка. Связанные друг с другом гомологичные хромосомы образуют хиазмы.
8. Хромосомное определение пола. Доза гена и тельца Барра.
XX – cамка (гомогаметный пол), ХY – самец (гетерогаметный пол)
Аутосомы – хромосомы одинаковые для обоих полов, половые - пара хромосом, различных у особей мужского и женского пола.
Половое размножение увеличивает генетическую вариабельность, то есть мейоз поддерживает генетическое постоянство вида.
Определение пола: генное (у гермафродитов, пол часто меняется) к хромосомному (раздельнополые, половыми хромосомами) к геномному (пчелы, муравьи. Самки из оплодотворенных яиц, самцы из неоплодотворенных. Плодовитость зависит от условий выкармливания).
Одновременно развивается половой диморфизм.
Средовое определение (нехромосомное) внешними факторами – у рептилий в зависимости от температуры. Партеногенетические поколения копепод.
Человек, позвоночные, насекомые и беспозвоночные: XX – cамка (гомогаметный пол), ХY – самец (гетерогаметный пол)
Некоторые насекомые: ХО – мужской, ХХ – женский
Птицы: ZZ – самцы, ZW - самки
Голандрический тип наследования - сцепленное с Y-хромосомой наследование. Гипертрихоз ушных раковин, избыточный рост волос на средних фалангах пальцев кистей
Полисомия по половым хромосомам — это состояние, при котором в клетках человека присутствуют лишние половые хромосомы. По Y мужчин это может быть 47,XYY, 48,XYYY, 48,XXYY или 49,XXYYY. У женщин — 47,XXX, 48,XXXX или 49,XXXXX. Синдром Дауна (трисомия по 21 хромосоме), синдром Клайнфельтера — полисомия по X-хромосомам у мальчиков
Тельце Барра (X-половой хроматин) — свёрнутая в плотную (гетерохроматиновую) структуру неактивная X-хромосома, наблюдаемая в интерфазных ядрах соматических клеток самок плацентарных млекопитающих, включая человека. У пробандов с синдромом Тернера (45, Х) их нет, при синдроме Клайнфельтера у мужчин (47, ХХY) обнаруживается одно, при 47, ХХХY кариотипе – 2.
Доза гена — это число копий определённого гена, присутствующих в геноме. В норме доза гена равна уровню плоидности хромосом, на которых ген локализован, однако может быть выше (трисомия и т. д.) и ниже (моносомия, половые хромосомы у гетерогаметного пола и т. п.) этого уровня.
9. Сцепленные признаки и их наследование. Кроссинговер и рекомбинация ДНК.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме.
Виды сцепленного наследования:
Полное сцепление. Гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. При скрещивании рецессивной по обоим признакам самки, имеющей тёмное тело и короткие крылья, с дигетерозиготным доминантным самцом дрозофилы образовалось 50% серых мух с длинными крыльями и 50 % мух с темным телом и короткими крыльями.
Неполное сцепление. Гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними. Если скрещивают дигибридную самку с гомозиготным рецессивным самцом, то в результате образуется потомство: 41,5 % — серых с длинными крыльями, 41,5 % — серых с короткими крыльями, 8,5 % — тёмных с длинными крыльями, 8,5 % — тёмных с короткими крыльями.
Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом.
Реципрокные скрещивания - совокупность двух скрещиваний с разной комбинацией родительских форм. Одна и та же форма выступает в одном скрещивании в качестве женского родителя, в другом — в качестве мужского.
Нерасхождение Х-хромосом — это случай, когда Х-хромосомы не расходятся в мейозе к разным полюсам, а направляются вместе к одному из полюсов. В результате этого образуются яйцеклетки с двумя Х-хромосомами и яйцеклетки без Х-хромосом. Трисомия по Х-хромосоме - в кариотипе присутствует дополнительная женская половая хромосома
Х-сцепленное наследование — это тип наследования, при котором признак передается через Х-хромосому. При этом поражаются мужчины, женщины могут быть носителями, но обычно здоровы; Примеры Х-сцепленных рецессивных заболеваний: дальтонизм (неразличение красного и зелёного цветов), гемофилия А и В.
Х-сцепленное доминантное наследование: пораженные мужчины передают заболевание только своим дочерям, а передача от отца к сыну невозможна.
Кроссинговер — это обмен участками гомологичных хромосом во время конъюгации в мейозе, приводит к рекомбинации сцепленных признаков или генов.
Конъюгация — это перенос генетического материала между двумя клетками посредством прямого контакта.
Рекомбинация ДНК — процесс обмена участками молекул ДНК путем разрыва и соединения разных молекул. Значение рекомбинации:
увеличение генетического разнообразия потомства — создание новых комбинаций аллелей;
репарация повреждений ДНК (восстановление);
ускорение эволюции.
*Дальше идет то, что есть в презентации, но вроде как не особо нужно в вопросе, поэтому пусть тоже будет на всякий случай
Сцепленные гены — это гены, наследуемые совместно в одной хромосоме. У человека — гены полидактилии (число пальцев выше нормы) и катаракты; у дрозофилы — гены цвета тела и длины крыльев.
Кроссинговер проходит по механизму гомологичной рекомбинации, предложен Холидеем - однонитевой разрыв в каждой из двунитевых молекул ДНК, вытеснение и замещение нити, миграция разрыва и разрешение единичной структуры Холидея.
Генная конверсия – последствие реципрокной рекомбинации (обмен участками ДНК между соответствующими сайтами на гомологичных хромосомах), при которой один аллель как бы превращается в другой. Возникает в результате формирования гетеродуплексов, содержащих неспаренные основания. После разрешения гетеродуплексов, исправления ошибок спаривания случайным образом иногда приводит к генной конверсии.
10. Рекомбинационные генетические карты.
Рекомбинационные генетические карты — это карты, которые показывают позиции генов и других последовательностей в геноме на основе определения частот рекомбинаций между генами.
Сантиморганида (сМ) — единица измерения генетической сцепленности между полиморфными фрагментами генома (локусами или маркерами). Она определяется как дистанция, на которой вероятность рекомбинации генов в мейозе составляет 1%.
Чем больше дистанция в сантиморганидах, тем меньше сцепленность участков ДНК, то есть больше вероятность кроссинговера этих участков.
Общая длина генетической карты организма рассчитывается как сумма минимальных экспериментально определенных генетических расстояний между парами маркеров. Для каждого вида эта величина своя, так как зависит от количества фенотипических маркеров, необходимых для построения полной генетической карты.
11. Фенотипическая и генотипическая изменчивость. Понятие мутации.
Фенотипическая изменчивость — это вариант изменений фенотипа, при котором генотип не затрагивается и полученные изменения не передаются по наследству; результат реагирования организма на условия окружающей среды.
Норма реакции - способность генотипа формировать разные фенотипы в онтогенезе, в зависимости от условий среды; пределы изменчивости признака. Наследуется не признак, а способность организма (его генотипа) в результате взаимодействия с условиями среды давать определенный фенотип. Распределение нормы реакции выражается в виде Гауссовой прямой.
Генотипическая (наследственная) изменчивость — способность живого организма изменять свой генотип. Благодаря генотипической изменчивости особь может приобретать признаки, ранее не свойственные ее виду.
Виды наследственной изменчивости:
Мутационная изменчивость — случайные скачкообразные наследуемые изменения признаков. Пример: появление животных-альбиносов, растений с измененной формой листьев или необычной окраской лепестков.
Комбинативная изменчивость — возникновение у особей различий, обусловленных новым сочетанием родительских генов. Возникает при половом размножении. При этом признаки родительских особей случайным образом комбинируются в ряду поколений, повышая разнообразие организмов в популяции.
Мутация — стойкое изменение генома, которое может быть унаследовано потомками данной клетки или организма.
Мутационная теория Г. Де Фриза, основные положения: – Мутация является скачкообразным стойким качественным изменением; – Мутации разнонаправленные (полезные и вредные) и могут повторяться; – Выявляемость мутаций зависит от выборки изучаемых организмов.
12. Классификация мутаций.
По характеру изменения генотипа: - Генные (точечные) - Хромосомные - Геномные
По характеру изменения фенотипа: - Летальные - Морфологические - Физиологические - Биохимические - Поведенческие
По проявлению в гетерозиготе: - Доминантные - Рецессивные.
По условиям возникновения: - Спонтанные (возникающие без видимых причин); - Индуцированные (в результате воздействия);
По степени отклонения от нормального фенотипа (Г. Меллер): - Гипоморфные. Изменённые аллели действуют в том же направлении, что и аллель дикого типа, но дают ослабленный эффект. - Аморфные. Проявляются в фенотипе как потеря гена. - Антиморфные. Изменяют фенотип дикого типа на противоположный. - Неоморфные. Фенотип мутантов совершенно отличен от дикого. - Гиперморфные. У этих мутантов количество биохимического продукта резко увеличивается.
По локализации в клетке: - Ядерные - Цитоплазматические (мутации внеядерных генов)
По возможности наследования: - Генеративные, т.е. в половых клетках. - Соматические, наследуются в случае вегетативного размножения (пример - злокачественные опухоли).
Плейотропный эффект мутаций - множественное проявление одной мутации, например, ненормальное развитие хрящей у мышей приводит к ряду серьезных патологий (сужение просвета трахей, утолщение ребер, изменение носовых ходов, общая задержка развития), недостаточность синтеза коллагена у человека – сколиоз, ПМК, плоскостопие, переразгибание суставов и др.
Экспрессивность — степень (сила) влияния какого-то гена на количественный признак. Например, площадь, занимаемая пятнами, будет характеризовать экспрессивность гена пятнистости.
Пенетрантность — вероятность того, что ген будет иметь любые фенотипические проявления.
13. Генные мутации и хромосомные перестройки. Геномные мутации
Генные мутации — это результат изменения нуклеотидной последовательности в молекуле ДНК в пределах одного гена. Если такая мутация происходит в гене, это приводит к изменению последовательности иРНК. В результате синтезируется другой белок, а в организме изменяется какой-либо признак.
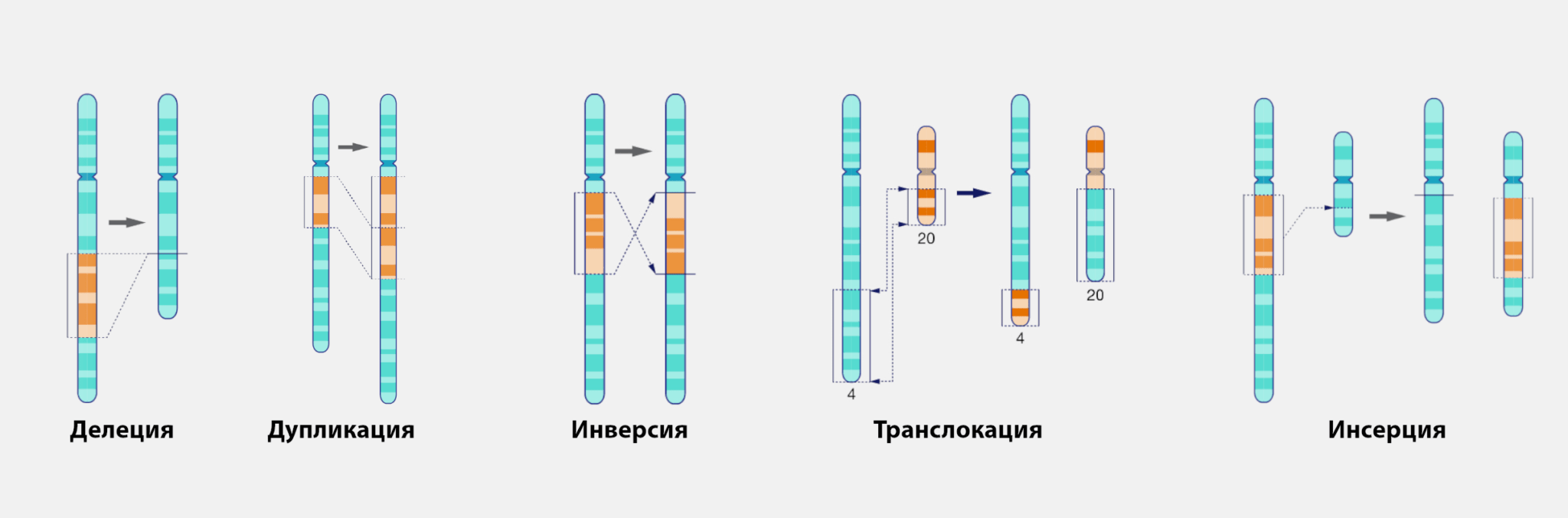
Хромосомные мутации — это изменения в структуре хромосом. Известны хромосомные перестройки разных типов:
делеция — выпадение участка хромосомы в средней её части;
дупликация — двух- или многократное повторение генов, локализованных в определённом участке хромосомы;
инверсия — поворот участка хромосомы на 180°, в результате чего в этом участке гены расположены в последовательности, обратной по сравнению с обычной;
транслокация — изменение положения какого-либо участка хромосомы в хромосомном наборе.
Геномные мутации — это изменения, которые возникают в результате изменения количества хромосом из-за сбоя в ходе мейоза или митоза.
Виды геномных мутаций:
Гаплоидия (также моноплоидия) — сокращение вдвое числа полных гаплоидных наборов хромосом.
Полиплоидия — увеличение количества полных гаплоидных наборов хромосом. В результате возникают триплоидные организмы (с набором 3n), тетраплоидные (4n) и т. д..
Гетероплоидия (также анеуплоидия) — некратное увеличение или сокращение числа хромосом. В большинстве случаев происходит увеличение или сокращение на одну хромосому
14. Типы полиморфизма геномов.
Полиморфизм геномов — это индивидуальные особенности геномов отдельных людей.
Основные типы полиморфизма:
Однонуклеотидный полиморфизм (ОНП). Отличия последовательности ДНК размером в один нуклеотид (А, Т, G или С) в геноме представителей одного вида или между гомологичными участками гомологичных хромосом индивида.
Полиморфизм инсерций-делеций ДНК. Результат изменений, вызываемых инсерцией или делецией от 2 до 100 нуклеотидов (инделов).
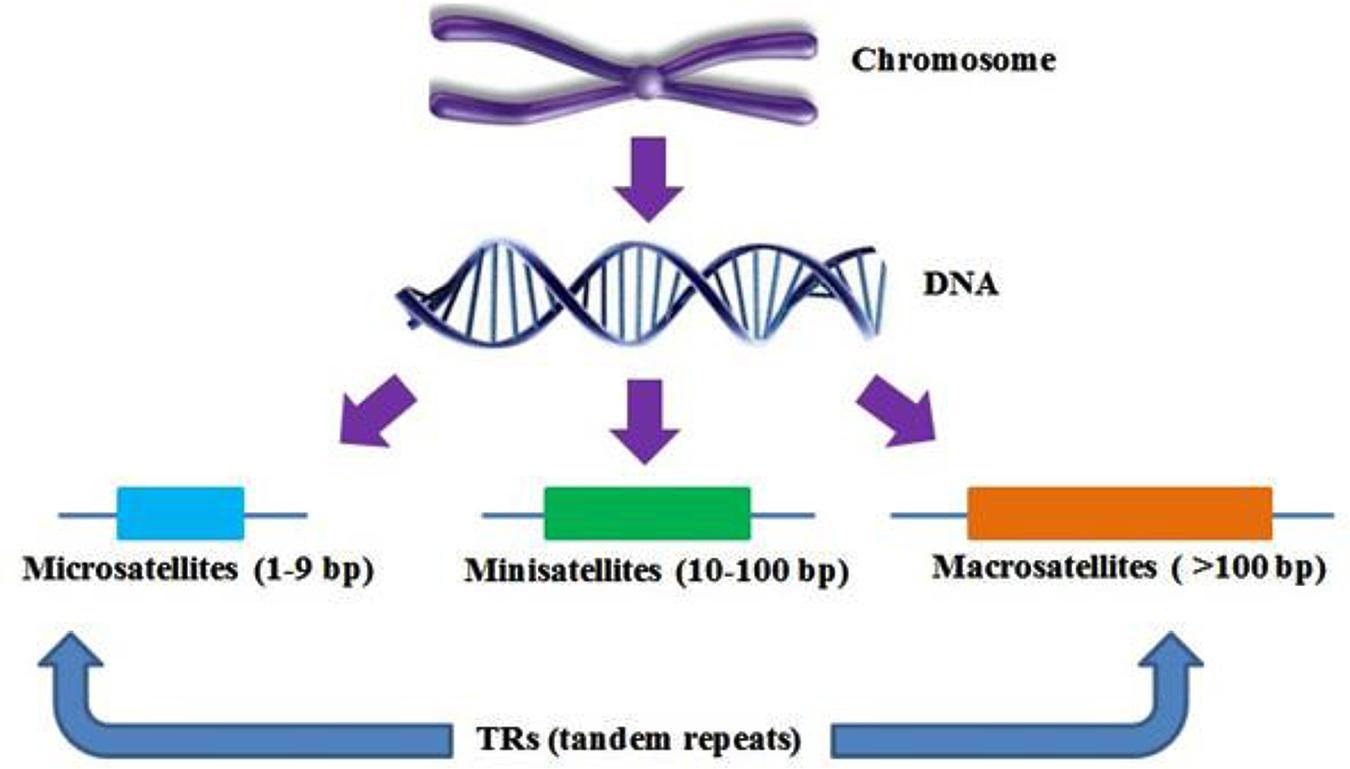
Микросателлитный полиморфизм. Разные аллели — результат различающегося числа повторов нуклеотидных блоков, содержащихся в микросателлите.
Мини-сателлитный полиморфизм. Этот класс полиморфизма имеет множество аллелей, различающихся числом тандемно повторяющихся копий мини-сателлита
15. Структура гена у про- и эукариот.
Общий план строения генов у прокариот и эукариот не отличается — и те, и другие содержат регуляторную область с промотором и оператором, единицу транскрипции с кодирующей и нетранслируемыми последовательностями и терминатор.
Особенности строения генов у прокариот:
Для прокариот характерно объединение нескольких генов в единую функциональную единицу — оперон.
Во время синтеза РНК на матрице ДНК с синтезируемой молекулой РНК может сразу связываться рибосома. Таким образом, трансляция начинается ещё до завершения транскрипции.
Особенности строения генов у эукариот:
У эукариот практически не встречается объединение генов в опероны.
Кодирующая последовательность гена эукариот чаще всего разделена на транслируемые участки — экзоны, и нетранслируемые участки — интроны.
Ключевые различия в структуре генов между эукариотами и прокариотами отражают их различные механизмы транскрипции и трансляции.
16. Структура ДНК и РНК. Основные формы спирали ДНК, их функциональное значение.
Нуклеотид — группа органических соединений, представляющих собой фосфорные эфиры нуклеозидов. Состоит из азотистого основания, пентозы (рибозы или дезоксирибозы) и остатка фосфорной кислоты.
Нуклеозид — фрагмент нуклеотида, который состоит из азотистого основания и моносахаридного остатка, соединённые N-гликозидной связью. Например, аденозин включает аденин, тимидин — тимин, уридин — урацил.
Структура молекулы ДНК — это закрученная по винтовой линии макромолекула, состоящая из двух антипараллельных полинуклеотидных цепей, стабилизированных межцепочечными водородными связями между специфическими парами азотистых оснований.
Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы — снаружи. Две спирали удерживаются вместе водородными связями между парами оснований.
Нуклеотиды двух цепей ДНК соединены водородными связями по принципу комплементарности: А-T, G-C. В паре A-Т 2H-связи, в паре Г-Ц – 3.
В двойной спирали цепочки также связаны с помощью гидрофобных связей и стэкинга, которые не зависят от последовательности оснований ДНК.
Пиримидиновые основания - производные пиримидина: У, Т, Ц Пуриновые основания - производные пурина: А, Г
Основные формы спирали ДНК:
A-форма — правозакрученная спираль - на виток приходится 11 пар азотистых оснований. Хранение наследственной информации о структуре специфических для каждого организма белков.
B-форма - правозакрученная - на виток приходится 10 комплементарных пар. Обеспечение передачи наследственной информации от клетки к клетке, от организма к организму.
Z-форма — левая спираль - на виток приходится 12 пар нуклеотидов, не имеет бороздок, но имеет изломы. Участие в реализации генетической информации, то есть в процессе синтеза полипептидов.
РНК (рибонуклеиновая кислота) — биополимер, представляющий собой одну цепочку нуклеотидов. Каждый нуклеотид состоит из пятиуглеродного сахара — рибозы, остатка фосфорной кислоты и азотистого основания.
Структура РНК организована в три уровня:
Первичный уровень. Состоит из линейной последовательности нуклеотидов.
Средний уровень. РНК сворачивается сама в себя за счёт внутримолекулярного спаривания оснований. Вторичная структура — это форма, которую она приобретает при складывании: спираль, петля, шпилька, внутренняя петля, псевдоузел и т. д..
Третичный уровень. Хотя РНК по своей структуре не образует двойную спираль, как ДНК, она обычно образует одинарную спираль как третичную структуру, поскольку её атомы взаимодействуют с окружающим пространством.
РНК переносят информацию о последовательности аминокислот в белках от хромосом к месту их синтеза и участвуют в синтезе белков.
17. Типы РНК в клетке: рРНК, мРНК, тРНК, мяРНК. Отличия структур мРНК про- и эукариот.
Клетки содержат четыре типа РНК, которые выполняют различные функции.
1.Информационная, или матричная. РНК (ее обозначают мРНК) считывает и переносит генетическую информацию от ДНК, содержащейся в хромосомах, к рибосомам, где происходит синтез белка со строго определенной последовательностью аминокислот. ~5%
2.Транспортная РНК (тРНК) переносит аминокислоты к рибосомам, где они соединяются пептидной связью в определенной последовательности, которую задает мРНК. ~5%
3.Рибосомная РНК (рРНК) непосредственно участвует в синтезе белков в рибосомах. Рибосомы — это сложные надмолекулярные структуры, которые состоят из четырех рРНК и нескольких десятков белков. Фактически рибосомы — это фабрики по производству белков. ~ 90% Основная РНК по массе в клетке. Рибосома функц-на и без белков, но в меньшей степени.
4.Малые ядерные РНК: интерферирующие РНК (iRNA), малые РНК, выполняют регуляторные функции.
Все виды РНК синтезируются на двойной спирали ДНК. мРНК у прокариот во время транскрипции синтезируется сразу зрела, и на ней может сразу проходить трансляция (синтез белков). мРНК эукариотов не сразу синтезируется зрелой: сначала синтезируется преРНК, которая потом подвергается сплайсингу (созреванию; удаляются некодирующие участки – интроны, а оставшиеся экзоны сшиваются лигазами?). мРНК синтезируется и далее трансляция проходят в цитоплазме у прокариот, у эукариот транскрипция проходит в ядре, а трансляция в цитоплазме.
18. Этапы репликации ДНК. Типы репликации геномов.
Репликация ДНК — это процесс создания двух дочерних молекул ДНК на основе родительской молекулы ДНК. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение.
Этапы репликации:
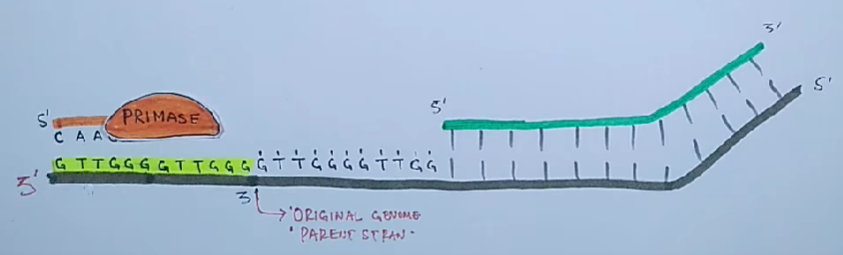
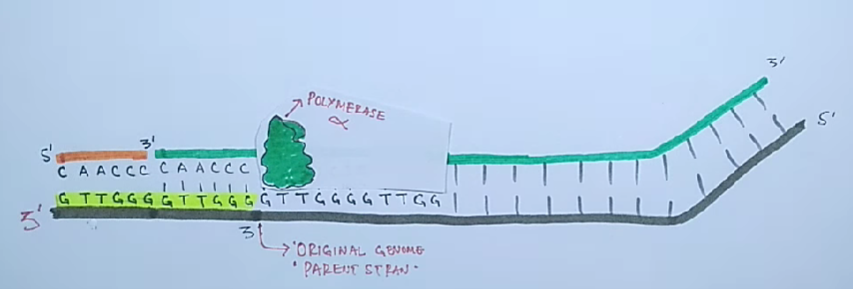
Инициация. Присоединение реплисомы к точке инициации /и деспирализация участка двойной спирали ДНК геликазами. Синтез РНК-праймеров ферментом праймазой, выполняющим функции РНК-полимеразы. Присоединение дезоксирибонуклеотидов, комплементарных матрице, к 3'-концу праймера ДНК-полимеразой.
Элонгация.Удлинение вновь синтезируемых цепей, которое осуществляется ДНК-полимеразой, входящей в состав реплисомы, во время её продвижения по матрице.
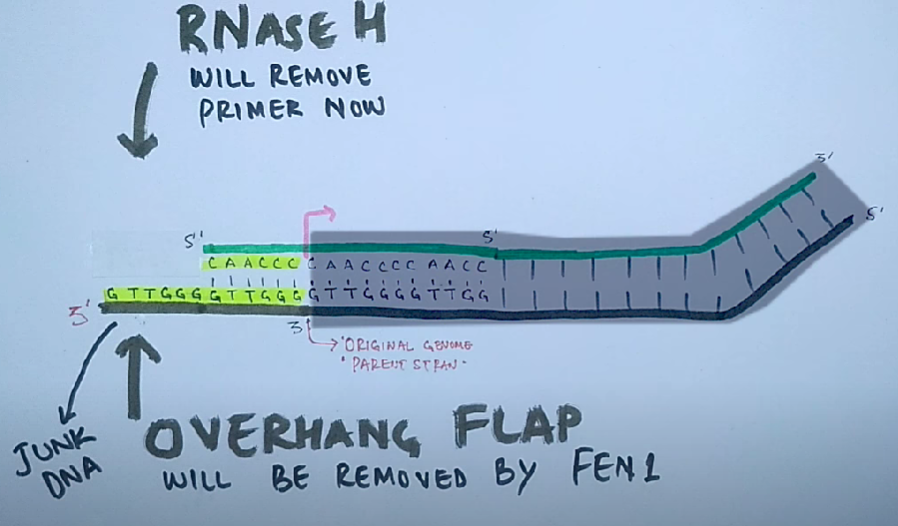
Терминация. Удаление РНК-праймеров 5'→3' экзонуклеазой, представляющей собой субъединицу ДНК-полимеразы. Заполнение недостающих участков ДНК-полимеразой. Связывание фрагментов вновь синтезированной ДНК с помощью лигаз.
Типы репликации:
Различают три типа репликации ДНК:
Полуконсервативный. Вновь синтезированная молекула ДНК состоит из одной материнской и одной дочерней полинуклеотидных цепей.
Консервативный. Вновь синтезированная молекула ДНК состоит только из дочерних полинуклеотидных последовательностей.
Дисперсный. Вновь синтезированная полинуклеотидная цепь ДНК состоит из фрагментов дочерних и материнских полинуклеотидных последовательностей.
Для прокариот и эукариот характерен полуконсервативный тип репликации. У вирусов встречаются все три типа репликации.
19. Проблема поддержание целостности линейной ДНК в ряду поколений. Теломеры и теломеразы.
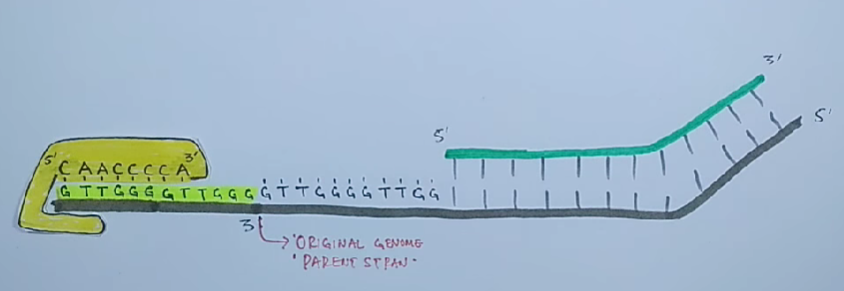
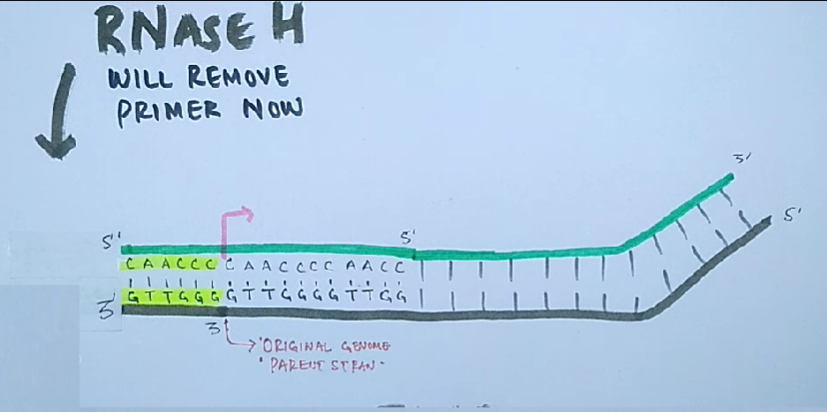
Проблема поддержания целостности линейной ДНК в ряду поколений заключается в том, что ДНК-полимеразы неспособны полностью реплицировать концы линейных молекул ДНК.
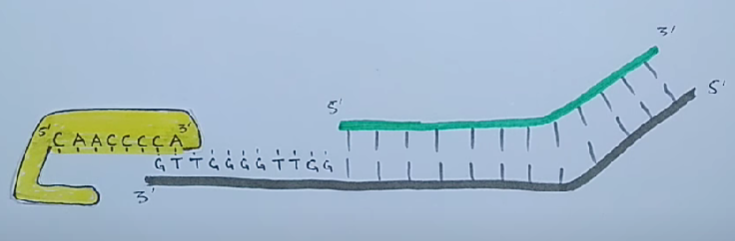
Для того чтобы клетки не теряли при делении часть генетического материала, 3'-концы ДНК хромосом эукариот наращиваются перед каждым раундом репликации короткими повторяющимися последовательностями. Это осуществляется ферментами — теломеразами.
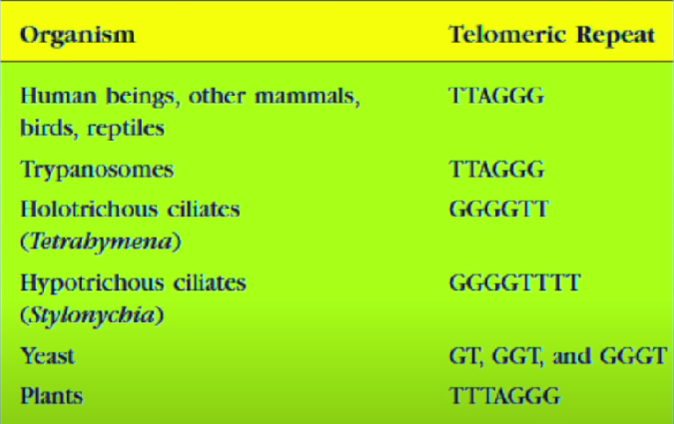
Теломеры — это области повторяющихся последовательностей на каждом конце хромосом большинства эукариот. Они защищают конец хромосомы от повреждения ДНК или от слияния с соседними хромосомами.
Теломераза — это фермент обратной транскриптазы, который несёт свою собственную молекулу РНК, которая используется в качестве матрицы при удлинении теломер. Теломераза активна в гаметах и большинстве раковых клеток, но в норме отсутствует в большинстве соматических клеток.
Износ теломер связывают со старением клеток, а их укорачивание объясняет тот факт, что каждая клетка способна делиться лишь определённое количество раз.
Стволовые клетки
имеют теломеразу. Теломераза в качестве
небелковой части (кофермента) имеют
простетическую олигонуклеотидную
группу, которая кодирует вырезанный
фрагмент - теломерные последовательности.
В соматических клетках недостроенная
последовательность удаляется
эндонуклеазой, поэтому с каждым делением
ген укорачивается.

20. Транскрипция и ее регуляция.
Транскрипция — это процесс копирования генетической информации из ДНК в РНК. Он проходит в три этапа:
Инициация. РНК-полимераза распознаёт промоторную область ДНК и связывается с ней. Двойная спираль ДНК разматывается, обнажая матричную нить для транскрипции.
Элонгация. Фермент перемещается вдоль цепочки ДНК-матрицы. По мере прогрессирования он синтезирует комплементарную цепь РНК путём включения рибонуклеотидов в соответствии с матрицей ДНК. Эта стадия продолжается до тех пор, пока не будет получен сигнал терминации.
Терминация. Транскрипция завершается, когда РНК-полимераза достигает специфического сигнала терминации ДНК. Этот сигнал даёт указание полимеразе высвободить новообразованную молекулу РНК. Двойная спираль ДНК реформируется, и продукт РНК высвобождается.
У прокариот регуляция транскрипции осуществляется на уровне изменения скорости синтеза мРНК. В основе теории лежит оперон — группа тесно связанных между собой генов, которые программируют образование структурных белков и ферментов в клетке.
У эукариот регуляция транскрипции осуществляется на двух взаимосвязанных уровнях:
На уровне аппарата и факторов транскрипции. Факторы транскрипции — это белки, обладающие способностью стимулировать или подавлять транскрипцию генов при связывании с регуляторными участками ДНК.
На уровне структуры хроматина.
От качества и
активности транскрипции зависит объём
синтеза тех или иных белков, жизнедеятельность
клетки, её способность адаптироваться
к окружающей обстановке

21. Созревание (процессинг) мРНК. Альтернативный сплайсинг.
Процессинг РНК — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК.
Основные этапы процессинга мРНК:
Кэппинг. К 5'-концу молекулы пре-мРНК добавляется модифицированный нуклеотид гуанин, который помогает защитить мРНК от деградации, а также необходим для эффективной трансляции.
Сплайсинг. Интроны (некодирующие области пре-мРНК) удаляются, а экзоны (кодирующие области) соединяются вместе, образуя зрелую молекулу мРНК. Этот процесс осуществляется комплексом, называемым сплайсосомой, который распознаёт определённые последовательности на границах интронов и экзонов.
Полиаденилирование. К 3'-концу молекулы мРНК добавляется длинная цепочка адениновых нуклеотидов. Этот поли(А) хвост помогает стабилизировать мРНК, а также участвует в регуляции её трансляции.
В результате процессинга образуется зрелая матричная РНК. В ядре клетки она взаимодействует со специальными белками и с их помощью через поры в ядерной мембране переносится в цитоплазму.
Альтернативный сплайсинг — вариант сплайсинга мРНК, при котором в ходе экспрессии гена (процесс, в ходе которого наследственная информация от гена преобразуется в РНК/белок) на основе одного и того же первичного транскрипта (пре-мРНК) происходит образование нескольких зрелых мРНК.
Наиболее распространённая разновидность альтернативного сплайсинга предусматривает пропуск экзона: отдельные экзоны транскрипта при определённых условиях могут быть как включены в зрелую мРНК, так и пропущены.
Белки, получаемые трансляцией таких мРНК, в результате имеют разные аминокислотные последовательности; таким образом, при альтернативном сплайсинге один транскрипт обеспечивает синтез нескольких белков.
Широкое распространение такого сплайсинга у эукариот приводит к значительному увеличению разнообразия белков, закодированных в их геномах. Например, организм человека синтезирует не менее чем 100 тысяч различных белков, в то время как число кодирующих их генов примерно 20 тысяч.
22. Генетический код и его свойства.
Генетический код — это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК или РНК.
Свойства генетического кода:
Триплетность. Каждая аминокислота кодируется тремя расположенными подряд нуклеотидами.
Универсальность. Генетический код един для всех живых организмов — от прокариот до человека.
Вырожденность (или избыточность). Одна и та же аминокислота может быть зашифрована несколькими триплетами (обычно от 2 до 6). Это делает хранение и передачу генетического кода более надежными. Лишь две аминокислоты (триптофан и метионин) являются исключением: они кодируются одним-единственным триплетом.
Однозначность. Каждый триплет может кодировать только одну аминокислоту.
Неперекрываемость. Один и тот же нуклеотид не может быть частью одновременно двух кодонов, расположенных рядом друг с другом.
Непрерывность – триплеты считываются один за одним без пропусков.
Специфичность – один триплет кодирует только одну аминокислот
Полярность – триплеты считываются в одном направлении
23. Вырожденность генетического кода и гипотеза качания тРНК.
Вырожденность генетического кода — это способность разных кодонов кодировать одну аминокислоту (кроме метионина и триптофана).
Эта способность не приводит к двусмысленности, так как для пары кодонов-синонимов, соответствующих одной аминокислоте, ни один из них не кодирует одновременно ещё какую-то аминокислоту. Чаще всего у таких кодонов совпадают первые две позиции.
Вырожденность предохраняет организм от генных мутаций: при замене одного нуклеотида в триплете есть вероятность, что будет синтезироваться всё та же аминокислота, и мутация не повлечёт за собой последствий для организма.
Гипотеза качания тРНК (гипотеза неоднозначного соответствия) объясняет способность тРНК узнавать более чем один кодон.
Эта гипотеза постулирует, что образование пары кодон-антикодон в двух первых положениях кодона происходит всегда по каноническим правилам, но в третьем положении возможно «качание» (неоднозначное соответствие). Объясняется это тем, что конформация антикодоновой петли тРНК допускает значительную подвижность первого основания антикодона.
Таким образом, одна и та же тРНК может узнавать несколько кодонов, кодирующих одну и ту же аминокислоту, но различающихся по одному нуклеотиду.
24. Трансляция и компоненты трансляционного механизма.
Трансляция — это процесс синтеза белка на основе информации, закодированной в молекуле РНК.
Основные участники процесса трансляции:
Рибосомы — комплексы белков и рРНК, которые обеспечивают место, где происходит синтез белка.
Трансляционные факторы — белки, которые помогают рибосомам в процессе трансляции, например, распознавая старт- и стоп-кодоны на РНК.
тРНК — переносят аминокислоты к рибосомам, где они добавляются к растущей цепи белка.
Процесс трансляции делится на три этапа:
Инициация. Малая субъединица рибосомы связывается с молекулой РНК, а трансляционный инициационный фактор помогает определить точное место начала синтеза белка.
Элонгация. Рибосома перемещается вдоль молекулы РНК, считывая триплеты нуклеотидов (кодоны) и связывая их с соответствующими аминокислотами.
Терминация. Рибосома достигает стоп-кодона на молекуле РНК, что сигнализирует о завершении синтеза белка.