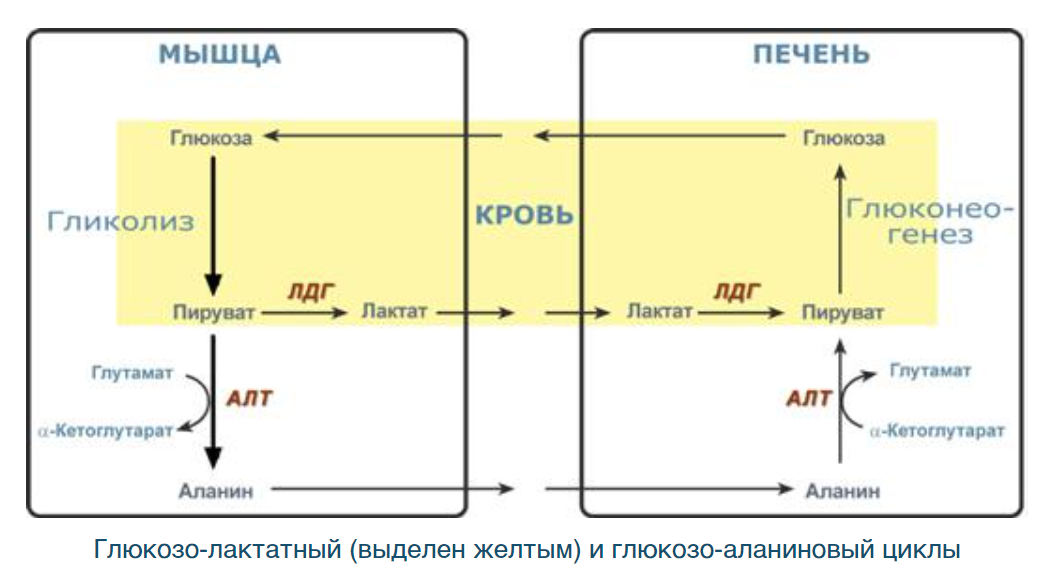
15. Глюкозо-лактатний цикл, схема взаємозв’язку гліколізу та глюконеогенезу. Роль лдг у м’язах та печінці. Гормональна регуляція процесів.
Целью глюкозо-аланинового цикла также является уборка пирувата, но кроме этого решается еще одна немаловажная задача – доставка аминного азота из мышцы в печень. При мышечной работе и в покое в миоците распадаются белки и образуемые аминокислоты трансаминируются с α-кетоглутаратом и полученный глутамат взаимодействует с пируватом. Образующийся аланин является транспортной формой аминного (аминокислотного) азота и пирувата из мышцы в печень. В гепатоците идет обратная реакция трансаминирования, аминогруппа через глутамат передается на синтез мочевины, пируват используется для синтеза глюкозы.
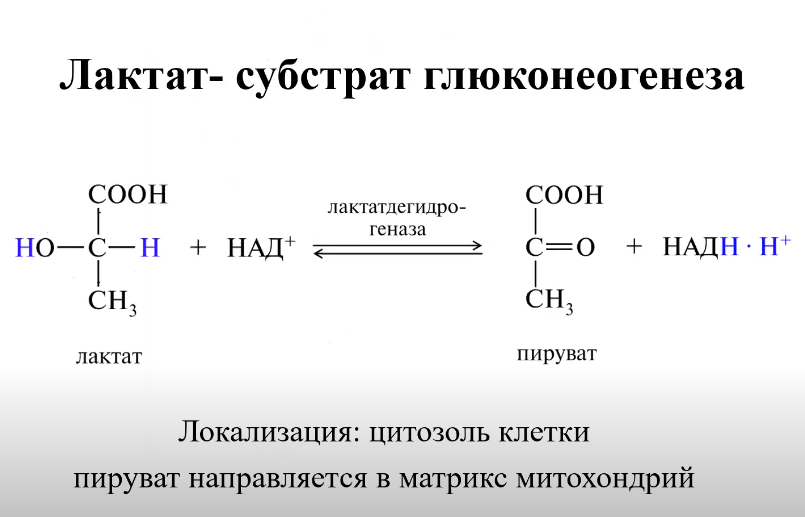
ЛДГ (лактатдегидрогеназа) – изоферменты, существует в виде 5 изоформ, каждая из которых состоит из 4-х субъединиц 2 типов М (muscle) и Н (heart):
ЛДГ1(НННН), ЛДГ2(НННМ): в тканях с аэробным обменом (миокард, мозг, корковый слой почек, МЫШЦЫ), обладают высоким сродством к молочной кислоте (лактату) и превращают его в пируват.
ЛДГ3(ННММ): в тканях с промежуточным типом обмена (селезенка, поджелудочная железа, надпочечники, лимфатические узлы)
ЛДГ4(НМММ), ЛДГ5(ММММ): в тканях, склонных к анаэробному обмену (ПЕЧЕНЬ, скелетные мышцы, кожа, мозговой слой почек), обладают низким сродством к лактату и катализируют превращение пирувата в лактат. Преобладают у зародыша.
Исследование уровня ЛДГ используется для первичной диагностики патологий в работе сердца, лёгких, печени, почек, некоторых онкологических заболеваний, а также деструкции мышечной ткани.
Основные гормоны, влияющие на глюкозо-лактатный цикл:
Инсулин стимулирует поглощение глюкозы мышцами и печенью, а также ингибирует глюконеогенез. Это приводит к накоплению пирувата в мышцах, который затем может быть превращен в лактат ЛДГ.
Глюкагон стимулирует глюконеогенез в печени, что приводит к превращению лактата, поступающего из мышц, в пируват. Пируват может затем использоваться для образования глюкозы.
Адреналин стимулирует распад гликогена в печени, что приводит к высвобождению глюкозы в кровь. Адреналин также стимулирует распад белков в мышцах, что может приводить к образованию аминокислот, которые могут использоваться для синтеза аланина.
Кортизол стимулирует глюконеогенез в печени и распад белков в мышцах.
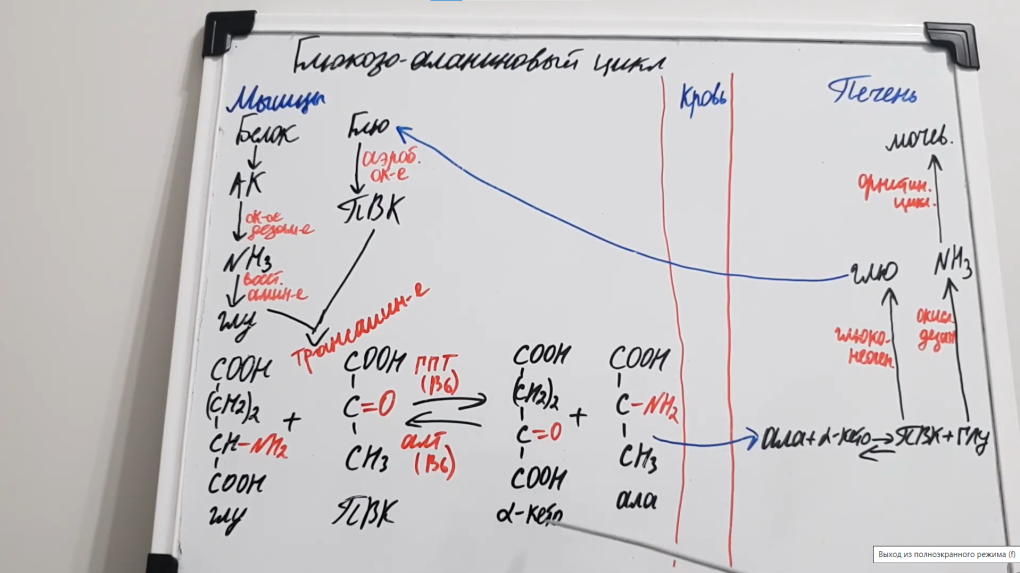
16. Глюкозо-аланіновий цикл, схема взаємозв’язку гліколізу та глюконеогенезу. Роль алт у м’язах та печінці. Гормональна регуляція процесів.
В мышцах основным акцептором лишнего аминного азота является пируват. При катаболизме белков в мышцах происходят реакции трансаминирования аминокислот, образуется глутамат, который далее передает аминоазот на пируват и образуется аланин. Из мышц с кровью аланин переносится в печень, где в обратной реакции передает свою аминогруппу на глутамат. Образующийся пируват используется как субстрат в реакциях синтеза глюкозы (глюконеогенез), а глутаминовая кислота дезаминируется и аммиак используется в синтезе мочевины.
Аланинаминотрансферазы, АЛТ, АлАТ — фермент печени, участвующий в обмене аминокислот. Наряду с АСТ, в большом количестве содержится АЛТ в печени, почках, в сердечной мышце, скелетной мускулатуре. При разрушении клеток этих органов, вызванных различными патологическими процессами, происходит выделение АЛТ в кровь человека, а анализ покажет высокий АлАТ в крови. В здоровом организме содержание показателя АЛТ в крови незначительно.
Биохимический анализ крови на АЛТ в крови покажет снижение уровня АлАТ при тяжелых заболеваниях печени — некроз, цирроз (при уменьшении количества клеток, синтезирующих АЛТ). Результаты анализа крови АлАТ покажет низкое содержание аланинаминотрансферазы при дефиците витамина В6. Повышение уровня АЛТ (альт аминотрансферазы) может быть признаком серьезных заболеваний, таких как: вирусный гепатит, токсическое поражение печени, цирроз печени, хронический алкоголизм, рак печени, токсическое действие на печень лекарств (антибиотиков и др.), желтуха, сердечная недостаточность, миокардит, панкреатит, инфаркт миокарда, шок, ожоги, травма и некроз скелетных мышц, обширные инфаркты, сердечная недостаточность.
Основные гормоны, влияющие на глюкозо-аланиновый цикл:
Инсулин снижает активность пируватдегидрогеназы, фермента, который катализирует превращение пирувата в ацетил-КоА. Это приводит к накоплению пирувата в мышцах, который затем может быть использован для образования аланина.
Глюкагон увеличивает активность пируватдегидрогеназы, что приводит к образованию ацетил-КоА в печени. Ацетил-КоА может использоваться для глюконеогенеза, процесса образования глюкозы из не углеводных субстратов.
Адреналин стимулирует высвобождение аминокислот из мышц, что обеспечивает субстратом для образования аланина.
Кортизол увеличивает активность глюконеогенеза в печени, что может привести к увеличению продукции аланина.
17. Молекулярно-клітинні механізми дії білково-пептидних гормонів та біогенних амінів. Адреналін, структура, синтез. Роль цАМФ як вторинного посередника в передачі гормонального сигналу. Процеси обміну вуглеводів та ліпідів, які регулюються адреналіном.
Структура: Гормоны надпочечников адреналин и норадреналин под общим названием катехоламины представляют собой производные аминокислоты тирозина. Роль адреналина является гормональной.
Синтез:

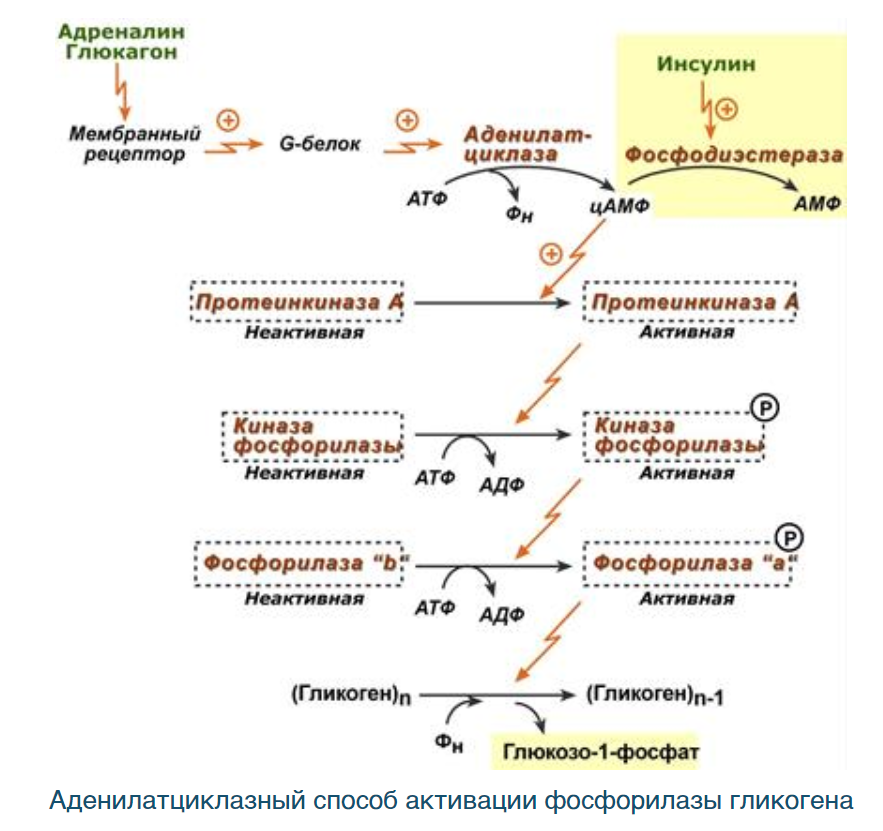
Механизм действия гормонов зависит от способности проникать их через плазматическую мембрану клетки. Водорастворимые гормоны белково-пептидной природы и адреналин не проходят через плазматическую мембрану, а взаимодействуют со специфическими мембранными рецепторами. В результате взаимодействия включаются внутриклеточные пути передачи информации, регулирующие метаболизм клетки и различные клеточные процессы. На уровне плазматической мембраны передача информации осуществляется путём последовательного изменения конформации мембранных белков (рецепторного, связующего) и фермента. Последний расположен с внутренней стороны мембраны и катализирует образование низкомолекулярного вещества – вторичного посредника, мессенджера. Диффузия вторичного посредника обеспечивает быстрое распространение сигнала по всей клетке к конкретным ферментам или другим белкам, реализующим ответ клетки на первичный сигнал – гормон или другое вещество (например, лекарства, бактериальный токсин), способные связываться с гормональным рецептором плазматической мембраны.
Непосредственной мишенью действия вторичных посредников служат ферменты протеинкиназы, которые путем фосфорилирования активируют или ингибируют специфические клеточные белки. Все составляющие компоненты составляют систему (каскад) и обеспечивают эффективную передачу и усиление соответствующего гормонального сигнала.
Первой была открыта аденилатциклазная мессенджерная система, в которой вторичным посредником является циклический АМФ. Структура цАМФ, реакция синтеза, которую катализирует мембраносвязанная аденилатциклаза, и реакция распада под действием фосфодиэстеразы рассмотрены ранее. Сигнал с гормональных рецепторов на аденилатциклазу передают G-белки двух типов: Gs-белок активирует аденилатциклазу, а Gi-белок тормозит. G-белки обоих типов состоят из альфа-, бета- и гамма-субъединиц, причем отличаются альфа-субъединицами, имеют центры связывания ГТФ и ГДФ и способны гидролизовать связанный ГТФ к ГДФ и неорганическому фосфату. До включения системы G-белок содержит связанный с альфа-субъединицей ГДФ и не взаимодействует с аденилатциклазой.
Присоединение гормона приводит к конформационным изменениям рецептора и G‑белка. Последний быстро связывает ГТФ вместо ГДФ и в такой форме изменяет активность аденилатциклазы (активирует или тормозит в зависимости от типа). Одновременно стимулируется ГТФазная активность G-белка и после перехода ГТФ в ГДФ активация аденилатциклазы прекращается. Белок-рецептор, G-белок и каталитическая субъединица аденилатциклазы вместе составляют аденилатциклазный комплекс. Трансмембранная передача сигнала комплексом завершается образованием цАМФ. Далее передача сигнала связана с действием ЦАМФ на внутриклеточные компоненты аденилатциклазной системы.
Работа аденилатциклазной системы подробно описывается при рассмотрении регуляции обмена гликогена, где адреналин запускает каскадный процесс активации гликогенфосфорилазы и инактивации гликогенсинтетазы.
18. Кортизол, синтез, механізм дії. Вплив на вуглеводний, ліпідний та амінокислотний обмін. Схема процесу глюконеогенеза. Включення в цю схему перетворень гліцерину та амінокислот. Вказати в схемі регуляторні ферменти та метаболічну регуляцію їх активності. Участь у цьому вітамінів.


Синтез: Глюкокортикоиды являются производными холестерола и имеют стероидную природу. Основным гормоном у человека является кортизол.
Механізм дії:
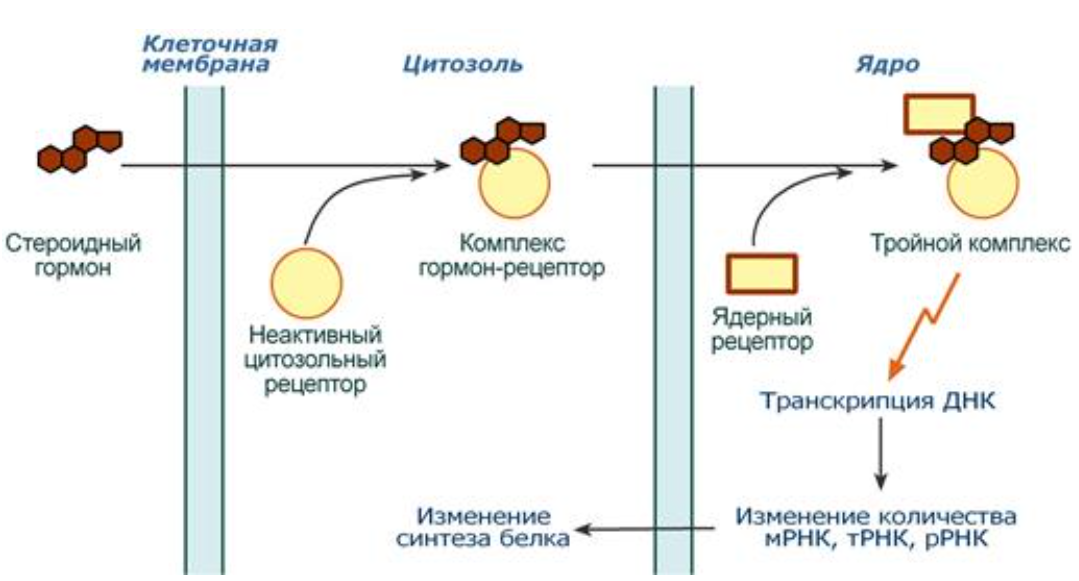
Схема процесу глюконеогенеза

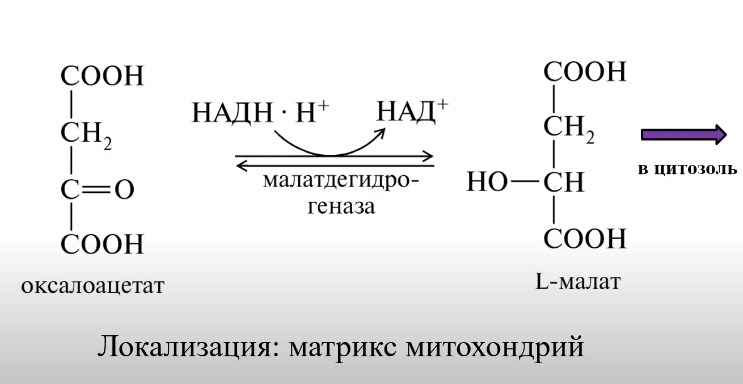
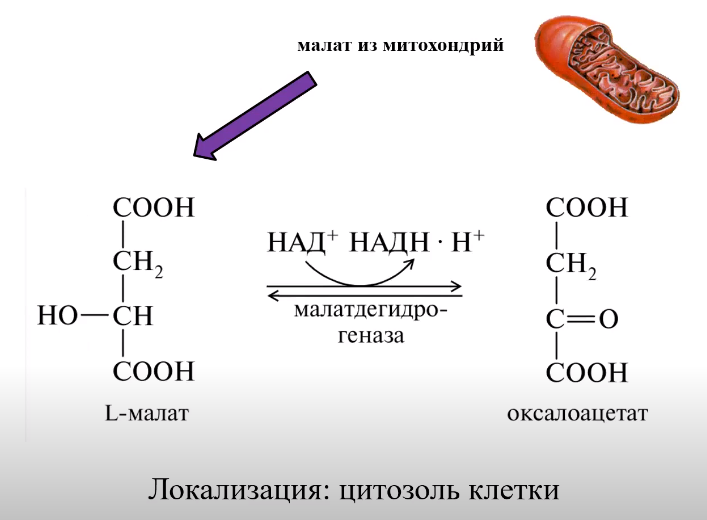
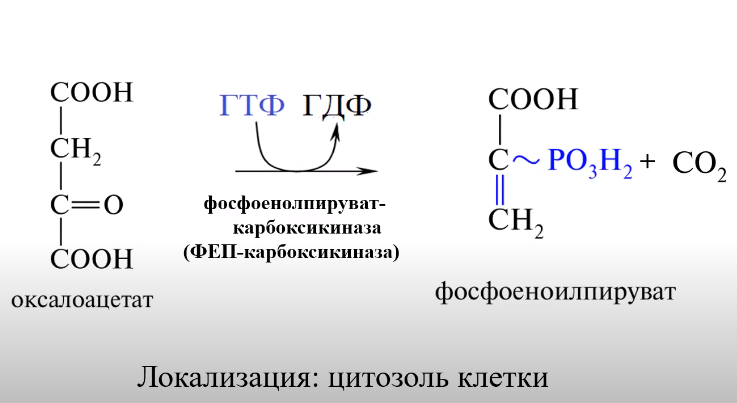
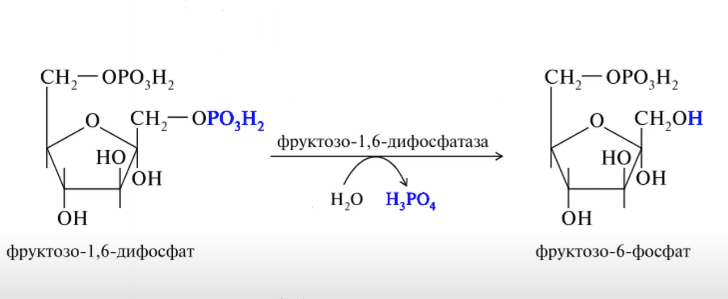
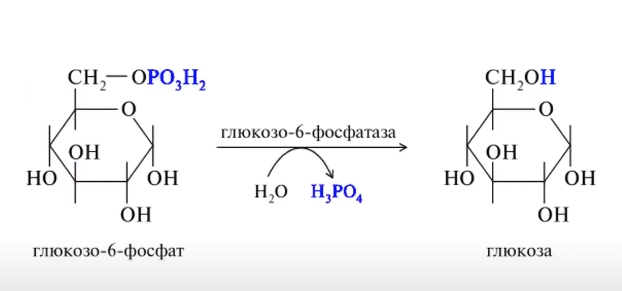
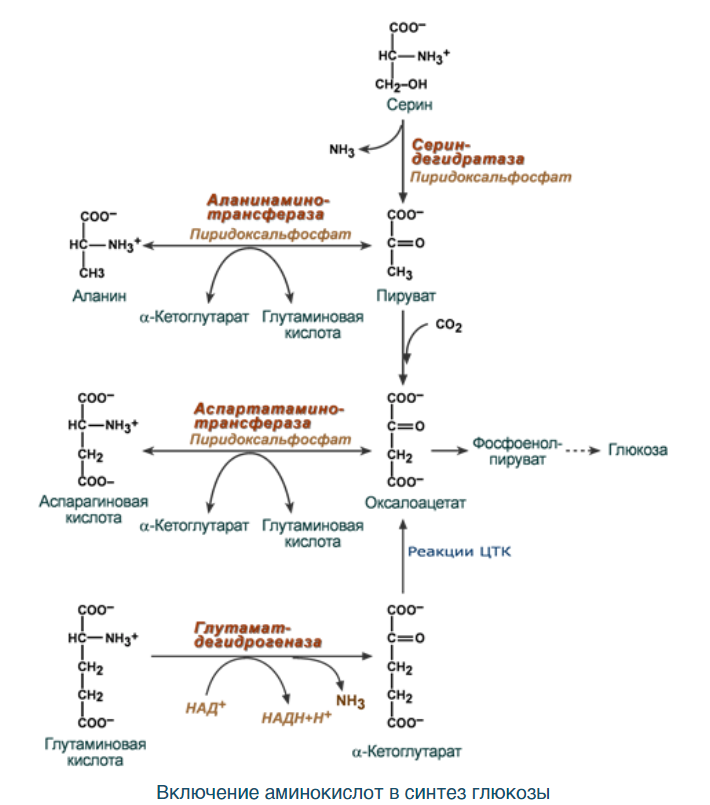
ВКЛЮЧЕНИЕ АК:
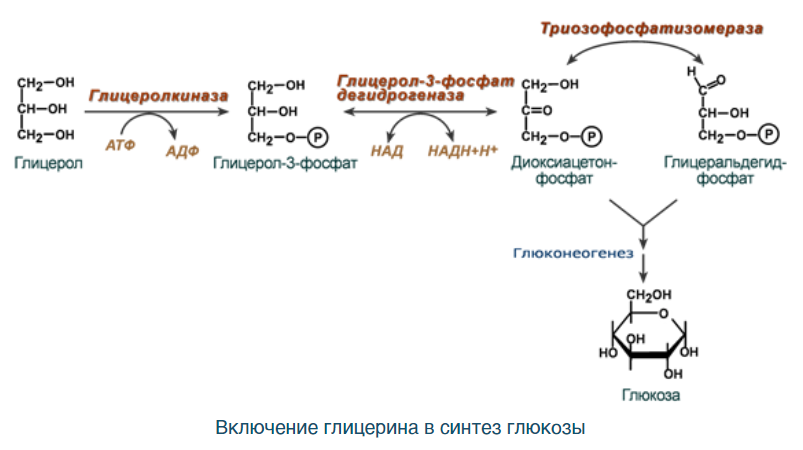
ВКЛЮЧЕНИЕ ГЛИЦЕРИНА:
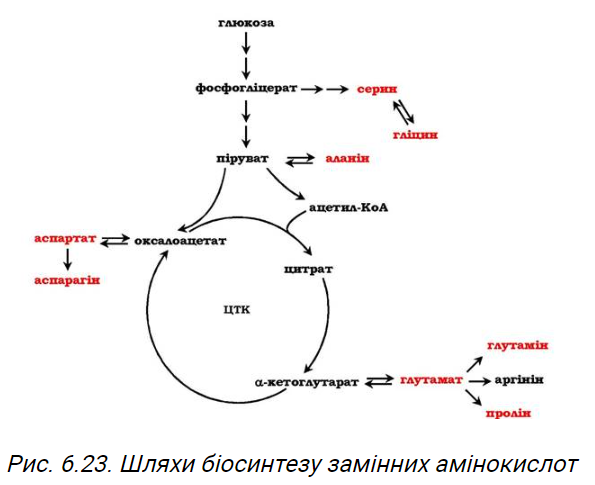
19. Пул вільних амінокислот в організмі: шляхи надходження та використання вільних амінокислот у тканинах. Синтез замінних амінокислот. Роль метаболітів ЦТК в цих процесах. Шляхи утворення та використання глютаміну в різних тканинах.
Пул свободных аминокислот (ПСА) - это динамическая совокупность свободных аминокислот, циркулирующих в крови и лимфе человека.
Источники свободных аминокислот: при переваривании белков из пищи аминокислоты всасываются в кровь, под действием протеолитических ферментов белки из мышц, кожи, других тканей и органов расщепляются на аминокислоты, некоторые аминокислоты (незаменимые) не могут синтезироваться в организме человека и должны поступать с пищей.
Синтез замінних амінокислот. Роль метаболітів ЦТК в цих процесах:
Метаболиты ЦТК – это промежуточные продукты цикла Кребса. Они играют важную роль в синтезе заменимых аминокислот, поскольку они могут быть превращены в кетокислоты, используемые для синтеза аминокислот.
Оксалоацетат, например, может использоваться для синтеза аспартата. Аспартат может быть аминатирован с участием глютамата, образуя глютамат, или может быть фосфорилирован, превращаясь в аспаргиновую кислоту. Глютамат, в свою очередь, может быть аминатирован с участием аммония, образуя глютамин. Таким образом, оксалоацетат через аспартат и глютамат помогает в синтезе аминокислот аспаргиновой кислоты и глютамина.
Альфа-кетоглутарат используется для синтеза глутамата, являющегося важным прекурсором для синтеза глутамина, аргинина и пролина. Глутамат может быть аминатирован с участием аммония, образуя глютамин. Глутамат также может быть конвертирован в аргинин и пролин.
Метаболиты
ЦТК, такие как оксалоацетат и
альфа-кетоглутарат, являются важными
интермедиатами для многих метаболических
путей, включая синтез заменимых
аминокислот. Они позволяют организму
эффективно использовать углеводы, жиры
и другие молекулы для синтеза необходимых
аминокислот.