

Механизм окислительного фосфорилирования и дыхательной цепи
Окислительное фосфорилирование – это многоэтапный процесс, происходящий во внутренней мембране митохондрий и заключающийся в окислении восстановленных эквивалентов (НАДН и ФАДН2) ферментами дыхательной цепи и сопровождающийся синтезом АТФ.
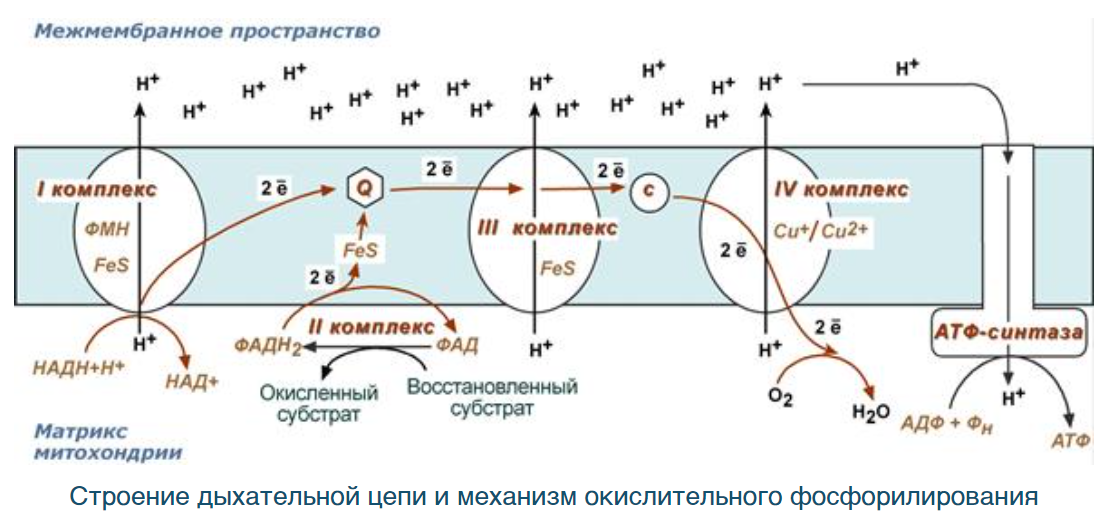
Пиридиновые дегидрогеназы содержат в качестве кофермента НАД. Субстраты малат, изоцитрат, пируват, глутамат окисляются НАД-зависимой дегидрогеназой, которая содержит кофермент НАД. Водороды от этих субстратов передаются на НАД, восстанавливая ее до НАДН2. Эта молекула идет к первому комплексу -- НАД-Н – дегидрогеназа, FMN которой забирает 1Н от НАДН2 и 1Н из воды в матриксе митохондрии, восстанавливаясь до FMNН2. Электроны от FMNН2 идут на FeS, а с него на убихинон, а протоны в межклеточное пространство с помощью СЭ. Перенос ионов водорода через мембрану (выкачивание) происходит в участках сопряжения. Они представлены I, III, IV комплексами дыхательных ферментов. В результате работы этих комплексов формируется градиент ионов водорода между внутренней и наружной поверхностями внутренней митохондриальной мембраны. Такой градиент обладает потенциальной энергией.
Второй комплекс (сукцинат-дегидрогеназа) содержит ФАД-зависимые дегидрогеназы, содержащие кофермент ФАД, и окисляющие сукцинат, оксалоацитат, ацил-КоА. Водороды от них переходят к ФАД = ФАДН2. Атомы отсюда сразу идут на убихинон.
Убихинон (отдельная структура) восстанавливается, превращаясь в убихинол. Электроны от него перейдут на 3 комплекс – убихинолдегидрогеназа.
С убихинола электроны передаются на 3 комплекс КоQ-цитохром c-оксидоредуктаза: б1, потом на б2, на FeS и наконец на с1. В каждом цитохроме происходит восстановление Fe3+ до Fe2+. Электроны передаются на цитохром с (отдельная структура), переносящий электроны на 4 комплекс -- цитохромоксидазный комплекс (цитохром а1 и а3, содержат гем и медь)
Электроны с него переноситься на О2, который связывается с протонами Н из матрикса митохондрии, образовывая метаболическую воду.
Свободная энергия с помощью, которой выбрасывались протоны, образовала электрохимический градиент; в матриксе -, снаружи +. Этот градиент нам нужен, чтобы протоны передались на АТФ-синтазу.
АТФ-синтаза содержит 2 части: F0 (канал, передающий протоны) и F1 (каталитическая -- синтез). Протоны тащат за собой энергию и градиент, запуская процесс превращения АДФ в АТФ, которая выброситься вон с помощью фермента транслоказа.

Роль о2
Роль кислорода: Кислород выступает в качестве акцептора электронов в реакции образования воды. В митохондриях кислород взаимодействует с электронами, возникающими во время электронного транспорта во время клеточного дыхания.
Окислительное фосфорилирование и его теория.
При прохождении электронов по дыхательной цепи происходит освобождение энергии. Однако благодаря большому количеству переносчиков выделение свободной энергии, которым сопровождается перенос одной пары электронов от НАДН2 на кислород, происходит не одномоментно, а разбивается на ряд относительно небольших порций. Поэтому дыхательную цепь можно рассматривать как каскад, при помощи которого клетка получает свободную энергию, извлекаемую из питательных веществ, в «расфасованном виде».
Коэффициент фосфорилирования это количество молекул АТФ, которое может быть синтезировано на одну молекулу NADH или FADH2 в процессе дыхательной цепи (окислительного фосфорилирования). Зависит от того, через какой комплекс дыхательной цепи электроны прошли
Общепризнанной теорией ОФ является хемиосмотическая (протондвижущая) теория П.Митчелла, по которой движущей силой ОФ является протонный потенциал (μН + ). Он возникает за счет того, что компоненты дыхательной цепи, присоединяя электроны, одновременно переносят протоны водорода в межмембранное пространство.
Таким образом в межмембранном пространстве на наружной поверхности внутренней мембраны митохондрии создается протонный (концентрационный) градиент. Градиент заставляет протоны двигаться из мембранного пространства внутрь в матрикс. Однако так как внутренняя мембрана митохондрий непроницаема для протонов, их движение в обратном направлении возможно только через особые каналы митохондриального белка F0 АТФсинтазы. Проходя через канал они изменяют конформацию белка F в результате чего фермент приобратает АТФсинтазную активность.
