

Физиология человека_Вопросы к экзамену_2024-1
.pdfТипы людей по Павлову:
Художественный тип (преобладает первая сигнальная система, восприимчивость к внешним раздражителям, образное мышление)
Мыслительный тип (преобладает вторая сигнальная система,
способность абстрагироваться от действительности)
Промежуточный (смешанный) тип (сбалансированность обеих сигнальных систем.
Функции речи: коммуникативная (общение между людьми), регулирующая
(побуждение к действию, волеизлияние, снижение порогов ощущения, регуляция поведения других людей), программирующая (предварительное построение схемы будущего высказывания и переходе этой схемы к воспроизведению высказывания).
Виды речи:
Устная:
-Монологическая (более сложное построение фраз)
-Диалогическая
Письменная (более точная)
Внешняя
Внутренняя:
обрывистая, понятная только самому человеку ! При переходе от внутренней к внешней – изменение структуры речи
Центры речи:
1.Центр Брока – задняя часть нижней (третьей) лобной извилины – моторный центр речи, отвечает за воспроизведение речи.
2.Центр Вернике – задняя треть верхней височной извилины - слуховой центр речи, отвечает за понимание услышанного.
3.Оптический центр речи – угловая (ангулярная) извилина – теменная

доля на границе с затылочной и височной – оптический центр речи, отвечает за понимание написанного.
Формирование речи:
Центры речи (располагаются в доминантном полушарии, например, для правшей (большинство населения Земли) – в левом полушарии)
Двигательные, зрительные, слуховые зоны коры
Пирамидная система
Экстрапирамидная система
Продолговатый мозг
Зрительный, глазодвигательный, лицевой, слуховой, языкоглоточный,
блуждающий, подъязычный нервы Восприятие речи – речеслуховой анализатор
Воспроизведение речи – речедвигательный анализатор
Нарушения функции речи:
Моторная афазия (поражение центра Брока) – невозможность правильно произносить слова
Сенсорная афазия (поражение центра Вернике) - непонимание слышимого
Оптическая афазия (поражение оптического центра речи) – неспособность распознавать написанное
Амнестическая афазия - поражение теменно-височной ассоциативной коры – нарушение функции подбора нужных слов
13.Нейрофизиологические основы сна. Метод электроэнцефалографии (ЭЭГ) и стадии сна. Нервные центры сна.
Ритмы ЭЭГ:
Основными анализируемыми параметрами ЭЭГ являются частота и
амплитуда волновой активности. Регистрируется четыре основных физиологических ритма: альфа-, бета-, тета- и дельта-ритмы .
В состоянии относительного покоя чаще всего регистрируется альфа-ритм (8 – 13 колебаний в 1 с);
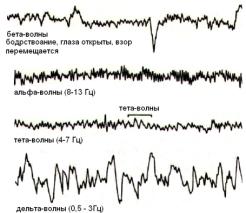
в состоянии активного внимания – бета-ритм (14 и выше колебаний в 1
с);
при засыпании и некоторых эмоциональных состояниях – тета-ритм (4
– 7 колебаний в 1 с);
при глубоком сне, потере сознания, наркозе – дельта-ритм (1 – 3 колебаний в 1 с).
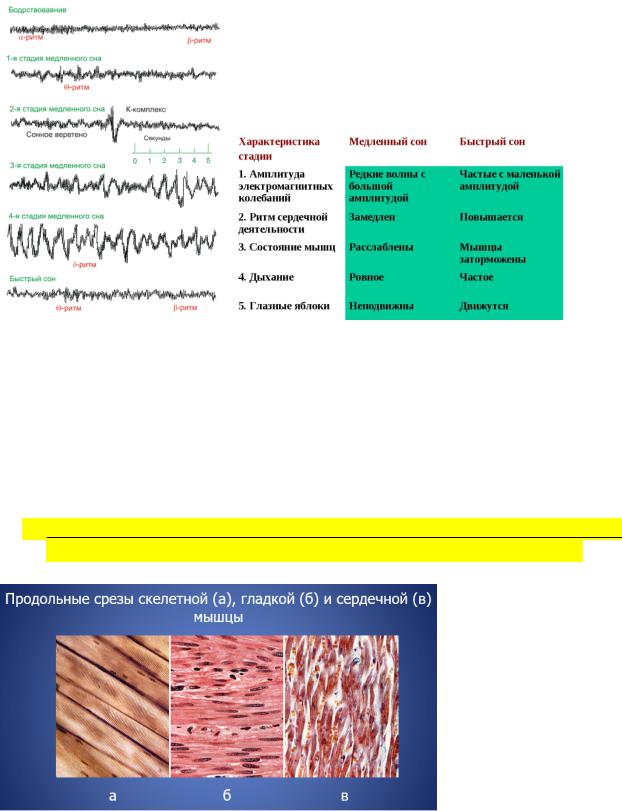
Стадии сна:
А - легкая сонливость, спокойное бодрствование (альфа-ритм - 8-13 колеб./сек; Гц, 30-70 мкВ)
В – нерегулярная активность (различная частота) – поверхностный сон
С – веретенообразные колебания (сигма-ритм, 12-14 колеб./сек)
D – смешанная, медленные высокоамплитудные колебания (дельта-ритм до 200-300 мкВ, 3-4 колеб./сек)
Е - медленные высокоамплитудные колебания (дельта-ритм 0,5-3 колеб./сек)
!Дельта-ритм – глубокий сон, наркотический сон, кома
Тетаритм (4-7 колеб./сек, 10-100 мкВ)
Р – парадоксальный сон, третье состояние; низкоамплитудные волны,
30 и > колеб./сек; длительность – 5-20 мин; 4-5 раз в течение ночи 4-6 циклов сна
Нервные центры сна:
1.Мост (голубое ядро)
2.Структуры зрительного анализатора
3.Кора б.п. (сформированные временные связи)
4.Гипоталамус и таламус при медленной стадии сна.
5.Голубое ядро, вестибулярные ядра, верхнее двухолмие среднего мозга, ретикулярная формация при быстрой стадии сна.
14.Структурная и функциональная организация
поперечно-полосатых (скелетных, сердечной) и гладких мышц.
Поперечнополосатая скелетная мышечная ткань:
•Имеет поперечнополосатую исчерченность, прикрепляется к скелету, приводит его в движение, а также способствует поддержанию равновесия тела в пространстве и является источником тепла.
•Может сокращаться быстро, со значительной силой.
Поперечнополосатая |
скелетная |
мышца |
состоит |
из |
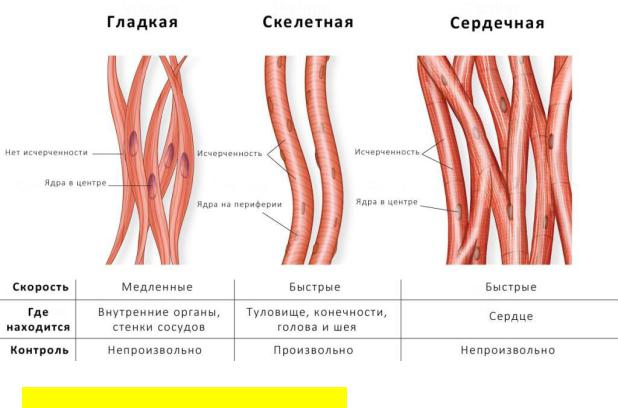
поперечно-исчерченных мышечных волокон, которые представляют собой многоядерные клетки (100 – 200 ядер).
Поперечнополосатая сердечная мышца:
•входит в состав сердечной стенки, имеет поперечнополосатую исчерченность, также способна к быстрому сокращению с большой силой.
•состоит из мышечных клеток (одноядерных) – кардиомиоцитов, формирующих сетчатое образованию, что способствует быстрому проведению возбуждения по мышце.
Гладкая мышечная ткань:
•входит в состав стенок внутренних органов, кровеносных сосудов, дыхательных путей, выводных протоков желез внешней секреции, имеет продольную исчерченность.
•практически непрерывно сокращается, однако с довольно низкой скоростью, то есть, способна к длительному, медленному тоническому сокращению.
•растяжение волокон вызывает быстрое сокращение ткани (такое явление называется перистальтикой), это способствует продвижению содержимого внутренних органов (например, перистальтика кишечника).
•структурной единицей гладкой мускулатуры является одноядерная клетка – миоцит.
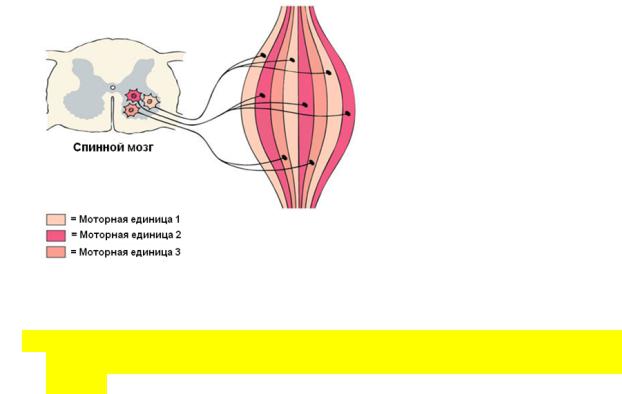
15.Понятие о двигательных единицах.
Двигательная единица – мотонейрон с группой иннервируемых им мышечных волокон.
16.Механизм сокращения и расслабления мышечного волокна
(поперечно-полосатая мышечная ткань и гладкая мышечная ткань).
1.Источником энергии для мышечного сокращения являются молекулы АТФ.
2.Гидролиз АТФ катализируется при мышечном сокращении миозином, обладающим ферментативной активностью.
3.Пусковым механизмом мышечного сокращения является повышение концентрации ионов кальция в саркоплазме миоцитов, вызываемое нервным двигательным импульсом.
4.Во время мышечного сокращения между тонкими и толстыми нитями миофибрилл возникают поперечные мостики или спайки.
5.Во время мышечного сокращения происходит скольжение тонких нитей вдоль толстых, что приводит к укорочению миофибрилл и всего мышечного волокна в целом.
В покоящейся мышце тонкие и толстые нити находятся в разъединенном состоянии.
Под воздействием нервного импульса ионы кальция выходят из цистерн саркоплазматической сети и присоединяются к белку тонких нитей – тропонину. Этот белок меняет свою конфигурацию и меняет конфигурацию актина. В результате образуется поперечный мостик между актином тонких нитей и миозином толстых нитей. При этом повышается АТФазная активность миозина. Миозин расщепляет АТФ и за счет выделившейся при этом энергии миозиновая головка подобно шарниру или веслу лодки поворачивается, что приводит к скольжению мышечных нитей навстречу друг другу.
Совершив поворот, мостики между нитями разрываются. АТФазная активность миозина резко снижается , прекращается гидролиз АТФ. Однако

при дальнейшем поступлении нервного импульса поперечные мостики вновь образуются, так как процесс, описанный выше, повторяется вновь.
17.Теория скольжения нитей. Роль саркоплазматического ретикулума
и ионов кальция в сокращении. Фазы мышечного сокращения. Режимы и виды мышечных сокращений. Энергетика мышечного сокращения. Способы ресинтеза АТФ, их мощность и емкость.
Теория скольжения нитей:
Теория скользящих нитей объясняет механизм мышечного сокращения, основанный на мышечных белках, которые скользят друг мимо друга, создавая движение.
Согласно теории скользящих нитей, миозин (толстые нити) мышечных волокон скользит мимо актина (тонкие нити) во время сокращения мышц, в то время как две группы нитей остаются относительно постоянной длины.
Роль саркоплазматического ретикулума и ионов кальция в сокращении:
В саркоплазматическом ретикулуме накапливаются ионы кальция, поступающие из саркоплазмы активным транспортом.
Главный интегральный белок мембраны саркоплазматического ретикулума выполняет функцию кальциевого насоса и при расслаблении мышцы перекачивает ионы кальция в ретикулум, используя для этого энергию АТФ.
К мышечному сокращению приводит повышение концентрации ионов кальция в саркоплазме и их присоединение к тропонину. Вследствие этого молекула тропонина изменяет свою конформацию и потому сдвигает тропомиозин в глубину желобка актиновой нити. Так на актиновой нити освобождаются участки, служащие местом прикрепления головок миозина.
Фазы мышечного сокращения:
1)Латентный период (напряжение 0)
2)Период сокращения (напряжение резко возрастает)
3)Период расслабления (напряжение постепенно уменьшается до 0)
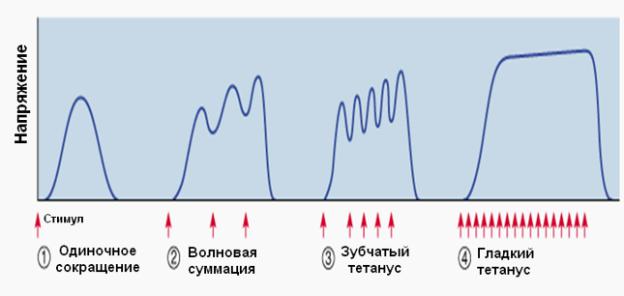
Виды мышечного сокращения:
Режимы мышечнного сокращения:
–изотонический режим – в данном случае выделяют два типа сокращения:
1)мышца находится в состоянии постоянного тонуса, мышца с одного конца закреплена, с другого конца свободно сокращается, нагрузка на
мышцу отсутствует (работа мышцы языка); 2) на мышцу оказывается нагрузка, при этом длина мышцы изменяется, а
напряжение остается постоянным (в чистом виде такой режим практически не встречается);
–изометрический режим – мышца находится в состоянии напряжения, при этом длина ее не изменяется (скольжения актиновых и миозиновых нитей не происходит), так как мышца закреплена с обоих концов (статическая работа);
–ауксотонический (смешанный) режим – длина и напряжение мышцы изменяются, мышца сокращается (выполнение динамической работы.)
Энергетика мышечного сокращения:
•АТФ – при расщеплении 1 моля АТФ выделяется 10 ккал энергии
•Креатинфосфат (взаимодействие АДФ и креатинфосфата с образованием АТФ и креатина) – 1 моль креатинфосфата = 10,5 ккал

энергии
•Анаэробный гликолиз – 1 моль глюкозы = 700 ккал энергии
•Аэробные реакции окисления углеводов и жиров – 1 моль жиров = 2400 ккал энергии
Способы ресинтеза АТФ их мощность и емкость:
15. Роль различных отделов ЦНС в регуляции движений.
Роль спинного мозга:
•Для поддержания мышечного тонуса достаточно рефлекторной деятельности спинного мозга, в этом случае импульсы к мышце поступают от альфа-мотонейронов спинного мозга.
•Двигательные рефлексы, за которые ответственен спинной мозг, осуществляются без участия сознанная, однако связь спинного мозга с головным является необходимой.
•В процессе регуляции рефлекторной деятельности со стороны отделов головного мозга задействованы гамма-мотонейроны спинного мозга.
1.Сухожильные рефлексы. Данные рефлексы возникают после нанесения удара молоточком по сухожилию, в результате чего мышца, прикрепленная к данному сухожилию, растягивается, в проприорецепторах возникает импульс (потенциал действия), который идет к спинному мозгу, от него – к мышце, вызывая ее сокращение.
2.Рефлексы растяжения. В случае быстрого растяжения мышцы, происходит ее сокращение (фазические рефлексы, примером являются сухожильные рефлексы). В случае медленного растяжения мышцы ее длина не изменяется (тонические рефлексы), что необходимо для поддержания равновесия тела в пространстве, так как мышца в этом случае сопротивляется растягивающей ее силе.
3.Рефлексы сгибания и разгибания. Рефлексы сгибания верхних и нижних конечностей наблюдаются после болевого раздражения последних. При этом на противоположной стороне тела часто происходит разгибание конечности. Данное явление связано с тем, что при активации мотонейронов мышц-сгибателей, через контрлатерали аксонов вставочных нейронов на противоположной стороне спинного мозга происходит активация мотонейронов мышц-разгибателей и торможение мотонейронов мышц-сгибателей. В некоторых случаях может иметь место одновременное возбуждение мотонейронов мышц-сгибателей и разгибателей.
4.Ритмические рефлексы. Наблюдаются при раздражении кожи (в случае, например, зуда), когда конечность совершает ритмические чесательные движения; при этом происходит чередование процессов сгибания и разгибание конечности. В данном случае сгибательный и разгибательный центры спинного мозга реципрокно тормозят друг друга. Ритмические рефлексы являются также основой шагательных рефлексов.
5.Рефлекс отталкивания от опоры (при ходьбе, беге).
6. Локомоция. Регуляция спинным мозгом координации движений, что является необходимым для перемещения тела в пространстве. В данном случае спинномозговые рефлекторые центры находятся под контролем ядерных образования ствола мозга, мозжечка, двигательной зоны коры больших полушарий вследствие влияния импульсов, поступающих от экстеро- и проприорецепторов.
Роль головного мозга:
•Тонус мышц – длительное напряжение мышцы, вызванное регулирующими влияниями нервной системы.
•Под влиянием ретикулярной формации мышечный тонус возрастает.
•Продолговатый мозг, наоборот, оказывает тормозящее влияние на тонус мышц. Причем, действие, как ретикулярной формации, так и ствола мозга в данном случае является неспецифическим, то есть распространяется на различные мышцы.
•Специфическое влияние различных отделов ЦНС заключается в их действии на отдельные группы мышц. В частности, кора больших полушарий через красные ядра среднего мозга
(кортикоруброспинальный тракт), ретикулярную формацию (ретикулоспинальный тракт), а также непосредственно через спинной мозг (короково-спинномозговой или кортикоспинальный тракт) усиливает тонус мышц-сгибателей, а продолговатый мозг посредством вестибулоспинального тракта усиливает тонус
мышц-разгибателей.
Голубое пятно моста угнетает мышечный тонус в фазу быстрого сна. Бледное ядро оказывает тормозящее влияние на мышечный тонус, а полосатое тело снимает это влияние.
Получая команды от вышележащих центров, красные ядра направляют импульсы по руброспинальному тракту к спинному мозгу и, таким образом, регулируют мышечный тонус.
Мозжечок участвует в поддержание мышечного тонуса: через красные ядра среднего мозга он активирует тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга – тонус мышц-разгибателей. Основную роль в поддержании мышечного тонуса, позы и равновесия тела играет кора червя.
Ретикулярная формация может оказывать тормозящее и активирующее влияние на осуществление ритмических рефлексов спинного мозга. Средний мозг участвует в регуляции движений (см. выше). Клетки черного вещества выделяют медиатор дофамин, который направляется к базальным ганглиям.
Автоматизирование движений и их содружественность находятся под контролем базальных ганглиев.
Считается, что базальные ганглии являются высшим эфферентным центром ствола головного мозга и регулируют двигательные безусловные рефлексы, а также вегетативные реакции, сопровождающие данные рефлексы.
Предполагается также, что базальные ганглии получают информацию от