

55 :: 56 :: 57 :: Содержание
12.3.9. Эволюция нефрона позвоночных
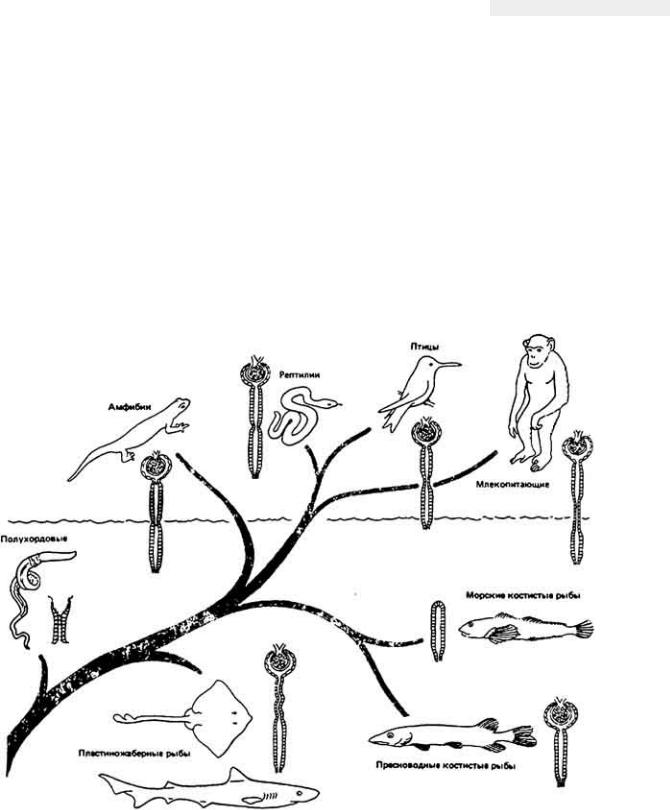
Схема эволюции нефрона позвоночных приведена на рис. 12-26. У некоторых современных полухордовых обнаружены примитивные предшественники почек с проксимальным концом нефрона, открытым во вторичную полость тела (целом). У ряда морских костистых рыб имеется простейший среди позвоночных животных нефрон, в котором отсутствуют как клубочек, так и боуменова капсула. В подобных бесклубочковых почках моча образуется полностью за счет секреции и, возможно, реабсорбции, потому что специального фильтрационного механизма нет. Противоположную крайность мы опять-таки видим
55
Рис. 12-26.
Упрощенная схема филогенеза почек позвоночных. (Smith, 1939.)
у морских животных, миксин (класс круглоротых); у них есть клубочки, но отсутствуют канальцы, и боуменовы капсулы впадают прямо в собирательную трубочку. Мы мало что знаем о физиологии таких нефронов, не считая их
способности активно выделять двухвалентные ионы Са2+, Mg2+ и SO 2- . Эти
4
почки не оказывают осморегулирующего эффекта, а если оказывают, то небольшой. Пропуская через себя в единицу времени некоторую часть крови, воды и кристаллоидов, они служат главным образом для профилактики накопления токсических отходов. В соответствии со сказанным внеклеточные

жидкости (табл. 12-2) наиболее примитивных из современных позвоночных, миксин, относительно сходны с морской водой по концентрации основных солей и их плазма по сути дела изотонична с морской водой.
Пресноводные костистые рыбы, как правило, имеют более крупные и более многочисленные почечные клубочки по сравнению со своими морскими родичами. Это справедливо и в отношении всех высших позвоночных. У низших позвоночных вплоть до рептилий почки не в состоянии выделять гипертоническую мочу (т.е. с большей осмотической концентрацией, чем в плазме крови). Способность почек продуцировать гипертоническую мочу четко коррелирует со структурной организацией нефрона и, если почка не имеет противоточного концентрирующего механизма, она не может выделять мочу с осмотической концентрацией заметно более высокой, чем в плазме. Как известно, только у млекопитающих, птиц и, возможно, пластиножаберных рыб почки обладают противоточной организацией, и поэтому только у данных животных выведение отходов, очевидно, основано на принципе осмотического противоточного повышения концентрации. Почки птиц похожи по строению на почки млекопитающих, если не считать, что у отдельных нефронов нет петли Генле, и у некоторых видов птиц петли ориентированы перпендикулярно к оси
56
собирательной трубочки, из-за чего снижается эффективность механизма концентрирования мочи. А у пластиножаберных рыб, например Raja erinacea (скат), было выявлено сложное строение почечных канальцев и наличие анатомических приспособлений для противоточного повышения концентрации выделяемой мочи. Однако нефроны ската в функциональном отношении совершенно иные, чем нефроны млекопитающих. Как мы уже видели, нефрон млекопитающих образует гипертоническую мочу за счет экскреции мочевины и задержки воды. Нефрон пластиножаберных, наоборот, задерживает мочевину, которая служит осмолитиком (см. разд. 12.1.2) и концентрированной мочи не образует. Таким образом, функция противоточного механизма нефрона пластиножаберных остается неясной.
57
55 :: 56 :: 57 :: Содержание

57 :: 58 :: 59 :: Содержание
12.4. Внепочечные саморегулирующие органы позвоночных
Хотя почки представляют собой наиболее сложные органы, выполняющие в организме функцию осмо-регуляции и экскреции, у позвоночных животных есть и другие органы, в равной степени важные для поддержания осмотического гомеостаза.
12.4.1. Осморегуляторная функция жабр у рыб
Чтобы жабры работали достаточно эффективно как орган дыхательного газообмена, они должны иметь большую эпителиальную поверхность (см. гл. 14). Хотя у таких животных, как рыбы, которые не находятся в состоянии осмотического равновесия с окружающей (водной) средой, эта особенность жабр создает для организма осмотические трудности, она же способствует исправному выполнению этими органами осморегуляторной функции. Поэтому у очень многих водных животных - позвоночных и беспозвоночных, - которые имеют жабры, последними осуществляется не только газообмен, но и такие различные функции, как перенос ионов, экскреция азотистых продуктов метаболизма и поддержание кислотно-щелочного равновесия. Например, у костистых рыб именно жабры выполняют центральную роль в преодолении осмотического стресса.
Строение жабр костистых рыб изображено на рис. 12-27, А. Кровь, омывающая жабры, течет по двум основным путям - приносящим (афферентным) и выносящим (эфферентным) сосудам, проходящим внутри жаберных нитей и через просвет внутри пластин. Эпителий, отделяющий кровь от внешней водной среды, состоит из двух различных типов клеток. Эпителий пластин целиком представлен плоскими клетками толщиной не более 3 - 5 мкм с небольшим числом митохондрий. Эти клетки, действуя как слабый барьер на пути диффузии газов, наилучшим способом обеспечивают дыхательный обмен. Эпителий, покрывающий жаберные нити, состоит из таких же клеток и клеток другого типа, которые по форме ближе к цилиндрическим и в несколько раз толще в направлении от основания к верхушке, чем плоские клетки. Их называют хлоридными клетками. На базальной и боковых поверхностях они имеют глубокие впячивания, образованные складками плазматической мембраны. Кроме того, указанные клетки очень богаты митохондриями и ферментами, обеспечивающими активный транспорт солей. В них содержатся в больших концентрациях глутаматдегидрогеназа и другие ферменты, связанные с обменом азотсодержащих веществ и экскрецией его конечных продуктов, а также карбоангидраза, необходимая для регуляции кислотно-щелочного состояния.
Хлоридные клетки впервые описали в 1932 г. Кейс (Ancel Keys) и Уилмер
(Edvard Willmer), считавшие, что они осуществляют транспорт хлоридов, поскольку эти клетки в гистохимическом отношении сходны с теми, которые в желудке амфибий секретируют соляную кислоту, и к тому же уже было известно, что жабры костистых рыб осуществляют выведение из организма хлорид-ионов (и натрия) без участия почек. Более современные гистохимические исследования подтвердили высокие уровни хлоридов в данных клетках, особенно вблизи ямки, возникающей на слизистой (наружной) поверхности их мембран, у рыб, которые адаптировались к высокой солености водной среды. Мы относительно мало знаем о том, как хлоридные клетки осуществляют транпорт солей. Однако научные данные позволяют считать, что хлориды выводятся из клетки не с помощью активного "выкачивания", а в результате пассивной утечки под влиянием разности потенциалов, возникающей между цитоплазмой и внеклеточной жидкостью вследствие активного переноса Na+ из клетки во внешнюю среду. Доказательством сказанного служит прекращение транспорта С1- в том случае, когда активность Na+ - К+ -АТРазы угнетается под действием специфического блокирующего вещества уабаина.
Интересно заметить, что у тех видов животных, которые мигрируют между соленой и пресной водой, направление переноса соли через эпителий жабр изменяется в зависимости от солености окружающей водной среды. В пресной воде организм активно поглощает соль, в соленой -экскретирует. В экспериментах, поставленных с целью изучения эффекта перевода таких рыб из воды с низкой концентрацией солей в воду с высокой соленостью, было установлено, что физиологическая адаптация жабр (т.е. переход от "накачивания" к "выкачиванию" NaCl) представляет собой постепенный
57
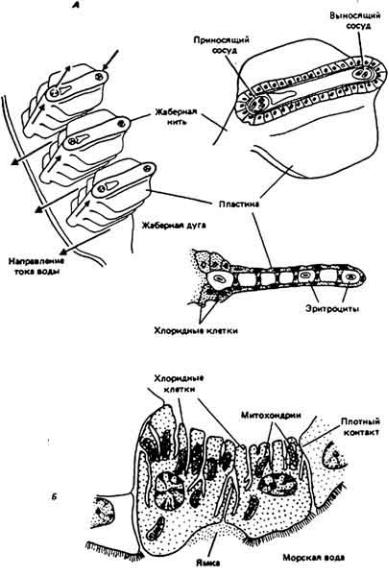
Рис. 12-27.
Жабры костистой рыбы как осморегуляторный орган. А. Жаберные нити выступают из жаберной дуги. Кровь течет по капиллярам в направлении, показанном цветными стрелками. Из нитей выступают плоские пластины (справа). В них содержатся обширные пространства, по которым циркулирует кровь. Б. Хлоридные клетки граничат с плоскими клетками дыхательного эпителия. Обратите внимание на изобилие митохондрий, а также многочисленные складки и щели, образуемые поверхностной мембраной клеток. (Maetz. 1971.)
процесс. Он включает в себя синтез и (или) разрушение молекулярных компонентов транспортной системы эпителия и изменения в морфологии и численности хлоридных клеток. Когда в эксперименте концентрацию соли в воде снижали, в хлоридных клетках происходили характерные изменения, включая исчезновение ямок. Воздействие раствора с высокой концентрацией соли, напротив, приводило к 1) появлению ямок, 2) увеличению численности хлоридных клеток, 3) соответствующему росту активности Na+-K+-ATPa3 и карбоангидразы, 4) расширению внеклеточных пространств за счет увеличения числа складок мембраны хлоридных клеток. Когда рыб, способных существовать при широком диапазоне концентраций соли в водной среде, например, лососевых, переносили из пресной в соленую воду, насос, "выкачивающий" ионы, начинал работать примерно через сутки. Ныне известно, что соленая вода вызывает указанные осморегуляторные реакции опосредованно через

эндокринные механизмы, которые влияют на дифференциацию и метаболизм эпителиальных клеток жабр. Стероидный гормон кортизол участвует в изменениях осмо-регуляции, обусловленных переводом рыбы из пресной в морскую воду, тогда как пролактин опосредует изменения, сопровождающие противоположный переход из соленой воды в пресную.
После того как хлоридные клетки впервые изучили и связали с транспортом С1-, было выявлено их участие в обмене других ионов, в том числе H+, Na+, К+,
NH + и HCO - . Трудно, конечно, точно доказать, что только хлоридные клетки и
43
никакие
58
другие участвуют в транспорте и обмене всех этих ионов. Однако многие факты, включая и те изменения, которым подвергаются хлоридные клетки при адаптации к высокой солености водной среды, свидетельствуют о главной роли данных клеток в процессе активного обмена ионов, происходящих в жабрах костистых рыб.
59
57 :: 58 :: 59 :: Содержание