Sekretarkiopkiopkiop18@yandex.rut.me/Prokururor I Вовсе не секретарь, но почту проверяю I steamcommunity.com/id/89885646844Опубликованный материал нарушает ваши авторские права? Сообщите нам.
жит основанием для исключения заявленного отцовства (при условии, что материнство рассматривается как бесспорное). В свою очередь полное комплементарное соответствие геномного профиля ребенка таковым заяв- ленных родителей означает неисключение заявленного родства.
Суммируя сказанное, в контексте судебно-экспертной идентификации личности результаты молекулярно-генетического типирования должны ин- терпретироваться в следующем логическом ключе.
А Сравнительный анализ: зафиксировано совпадение или несовпадение локальных генетических профилей либо (в случае экспертизы родст- ва) индивидуальных аллелей, выявленных у идентифицирующего объекта и у идентифицируемого лица, или соответственно у ребенка и родителей.
▲ Вероятностный анализ: в случае совпадения генетических профилей или аллелей — какова статистическая значимость этого события?
44.4.2. Сравнительный анализ геномных профилей
Формальное сравнение. В принципе два геномных профиля можно счи- тать одинаковыми (совпадающими), если они неразличимы по генотипу. Для индивидуализирующих систем, основанных на использовании поли- морфных локусов с дискретным распределением аллелей, это означает со-
впадение в сравниваемых геномных профилях всего набора составляющих аллелей. Чтобы это утверждать, надо, очевидно, уметь определить (иденти- фицировать) сами аллели. В этом есть свои особенности.
Дело в том, что прямое (безусловное) генотипирование, т.е. прямое ус- тановление локального генотипа, возможно только для систем, использую- щих аллельспецифичные диагностикумы, например ППАФ-типа.Так, при ППАФ-анализе локуса HLA DQA1 и локусов системы PolyMarkerR иденти- фикация аллелей легко достигается за счет того, что регистрируемые дот- сигналы являются аллельспецифичными и потому непосредственно указы- вают, какой именно аллельный вариант выявлен. Регистрируемые таким образом генетические характеристики имеют абсолютное значение и пото- му могут помещаться в банк данных для последующего сравнения с любы- ми другими аналогичными данными.
В тех же случаях, когда прямое генотипирование невозможно, необхо- димо соблюдать определенные условия, которые должны обеспечить кор- ректность процедуры генотипирования. Например, для ПДАФ-систем на основе гипервариабельных локусов с варьирующим числом тандемных по- второв, которые предполагают электрофоретическое фракционирование амплифицированных фрагментов ДНК, аллели определяют не прямым ме- тодом, а опосредованно — на основании физического расположения (пози- ционирования) фрагментов ДНК на дорожке геля. Но тогда для идентифи- кации аллелей необходимы правила, которые определяли бы критерии по- зиционного совпадения полос на геле. Важно понимать, что такие крите- рии различаются в зависимости от свойств аналитической системы и спо- соба сравнения (см. ниже). Поэтому регистрируемые генетические характе- ристики имеют не абсолютное, а относительное значение. Иначе говоря,
они могут быть пригодны для сравнения в рамках одного эксперимента и не могут использоваться для более широкого исследования. (Поэтому в банк данных они могут помещаться только тогда, когда установлено их со- ответствие такому уровню точности, который обеспечивает возможность сравнения с любыми аналогичными данными.)
515
33"
На практике совпадение геномных ПДАФ-профилей фиксируется в том) случае, когда картины распределения полос на сравниваемых дорож- ках! геля однотипны и похожи, а позиции соответствующих фрагментов попадают в интервалы, удовлетворяющие установленным допускам.
Подобные принципы сравнительного позиционного анализа примени- мы и для индивидуализирующих систем, которым свойственно непрерыв- ное распределение аллелей, например, таких как минисателлитные моно-и мультилокусные ПДРФ-системы,поскольку для них определение генотипа еще более проблематично. В этом случае существуют проблемы как теоре- тические, так и практические. Так, индивидуально-специфическая картина гибридизации, состоящая из множества характеристический полос и получающаяся при мультилокусной геномной дактилоскопии, представля- ет собой своего рода «черный ящик», в котором остаются не^ выясненны- ми ни генетическая природа, ни аллельные состояния локусов, соответст- вующих этим полосам. Таким образом, поскольку для мультило-кусных систем критерии генетического тождества полос не определены, по сути сравнение осуществляется не по самому генотипу, а как бы п<я внешнему образу генотипа. В данном случае это оказывается возможным благодаря
выраженной графической индивидуальности мультилокусного геномного профиля, который характеризуется сразу несколькими параметрами: чис- лом полос, их расположением на дорожке и интенсивностью каждой поло- сы.
При этом, однако, надо помнить, что оценочная специфичность муль-т тилокусного геномного «отпечатка» зависит от конкретных условий про- ведения анализа, параметров и особенностей экспериментальных проце- дур, количества и качества анализируемой ДНК. Это имеет принципиаль- ное значение, поскольку отождествляемые препараты ДНК могут значи- тельно различаться по своим «дактилоскопическим» свойствам, в частно- сти вследствие процессов деградации нативных молекул ДНК. Таким об- разом, при интерпретации результатов следует учитывать всю совокуп- ность экспериментальных условий, действующих факторов и получаемых данных. Следует признать, что пока для мультилокусных систем эта задача в общем виде решена не полностью, хотя определенный прогресс наблю- дается в решении] некоторых частных вопросов.
Интерпретация гибридизационной картины в случае использования мо-нолокусных зондов проще, чем для мультилокусных систем. Принци- пиальное значение приобретает тот факт, что теоретически в геномных профилях сравниваемых объектов совпадающие полосы заведомо тожде- ственны, поскольку они представляют один и тот же аллельный вариант конкретного генетического локуса, т.е. один и тот же индивидуализирую- щий признак. Следует, однако, оговориться, что в свою очередь для моно- локусных систем более проблематичным может оказаться решение вопро- са собственно совпадения полос из-за сплошного характера их распреде-
ления и отсутствия внутренних маркеров геометрической однородности геля. (В мультилокусных системах роль таких маркеров играют достаточ- но многочисленные мономорфные полосы.) Для решения этой проблемы предложены специальные методические и биостатистические подходы, в частности так называемый бининг (от англ. bin — интервал), когда аллели учитывают не индивидуально, а определенными группами.
В дальнейшем мы сосредоточим внимание на вопросах, связанных с
интерпретацией только амплификационных геномных профилей локусов с варьирующим числом тандемных повторов, поскольку в нашей стране именно они имеют наибольшее практическое значение.
516
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
Позиционное сопоставление амплификационных геномных профилей. Во- прос о том, одинаковы или неодинаковы геномные профили, которые по- лучены при анализе препаратов ДНК, выделенных из объектов экспертизы (например, из биологических следов на вещественных доказательствах и из крови проходящих по делу лиц), является ключевым для молекулярно-ге-нетической идентификации. Это ясно, поскольку от того, как будет интер- претирован результат сравнения, зависит экспертный вывод: в одном слу- чае это неисключение, а в другом —исключение причастности данного лица к происхождению исследованных следов. Между тем именно вопрос о
похожести и непохожести геномных профилей может создать большую опасность неверного решения из-за множества причин. Некоторые из них мы рассмотрим в данном разделе.
Итак, как уже упоминалось, при анализе электрофореграммы можно выделить два аспекта:
•картины распределения полос на сравниваемых дорожках геля похо- жи или непохожи;
•позиции соответствующих фрагментов совпадают или не совпадают.
Л о ж н о е г е н о т и п и р о в а н и е . Что касается общей похожести одного геномного профиля на другой, то в данном случае ложный результат может быть обусловлен двумя причинами.
Во-первых,присутствие в сравниваемых препаратах ДНК постороннего генетического материала (чаще всего в результате случайных загрязнений) может имитировать как совпадение, так и различие их геномных профилей. Во-вторых,этот же эффект может проявиться как результат неправильного генотипирования, в частности в результате ложноопределеинои гомо- или гетерозиготности анализируемых объектов. Это связано с артефактами ПЦР, возникающими под влиянием неоптимальных условий ее проведе- ния:
—избыточным или недостаточным исходным количеством матричной ДНК;
—плохим качеством препарата;
—неспецифичностью праймеров и(или) неадекватно подобранным для них рабочим режимом (например, отжиг при более низкой, чем сле- дует, температуре);
—неоптимальной концентрацией Taq-полимеразы в реакционной сме- си, в частности, ее избыточным количеством;
—присутствием в реакции неоптимальной концентрации Mg2+;
—профилем кривой нагрева реакционной смеси с неоптимальными значениями температурных переходов (это может быть вызвано не- удачными техническими параметрами прибора, используемого для амплификации ДНК);
—неоптимальной продолжительностью процесса циклического нара- щивания.
Наиболее распространенный артефакт, так называемый феномен пред- почтительной амплификации аллелей, довольно часто приводит к ошибоч- ному заключению о гомози готности. Вместе с тем наряду с опасностью ти-
пирования ложных гомозигот нередки и более сложные случаи искажения генотипа, характеризующиеся не только частичной утратой истинных алле- лей, но и амплификацией неспецифических (неаллельных) фрагментов, что имитирует ложногетерозиготныи аллельныи профиль. Решить проблему ложноопределеинои гомо- или гетерозиготности в некоторых случаях
517
сложно. Помочь в этом случае может только скрупулезный анализ устойчи- вости амплификационных профилей, основанный на амплификационнои титровании сомнительных препаратов и многократной проверке воспроиз- водимости результата.
С д в и г п о л о с . Физическое сопоставление полос на электрофоре- грамме также требует учета многих факторов. Одна из сложностей — тая называемый сдвиг полос, когда в процессе электрофореза фрагменты ДНК в одной дорожке геля движутся быстрее или медленнее, чем идентичны! фрагменты в соседней дорожке. Это явление имеет несколько причин:
—неодинаковое количество ДНК в разных дорожках геля;
—неодинаковые ионные характеристики препаратов, внесенных в раз-3ные дорожки геля (присутствие в препаратах примесей, влияющих на электрофоретическую подвижность ДНК — солей, спирта, фенола, этидиумбромида и др.);
—избыточная напряженность электрического поля;
—перегрев и нарушение структуры геля (избыточный ток);
—истощение электрофорезного буфера;
—микрогетерогенность геля (в частности, недостаточно высокое каче- ство среды разделения).
Большая часть перечисленных факторов поддается контролю, поэтому решением проблемы может быть оптимизация и текущий мониторинг соот-
ветствующих параметров.
В некоторых случаях сдвиг полос имеет не ступенчатый, а сглаженный характер (например, хорошо известный эффект «улыбки»). Тогда вознщ
кающие геометрические искажения электрофоретической картины в принт ципе поддаются математическому анализу и могут быть скомпенсированы на стадии обработки изображения. Для этого рекомендуется, кроме флан- кирующих дорожек, также и каждую 3—4-ю дорожку геля делать референт- ной, т.е. вносить в нее маркеры молекулярных масс, по которым будет рас- считываться фактор коррекции. Следует, однако, отметить, что надежная коррекция возможна не всегда и электрофорез по возможности надо повто-
рить.
Р а з р е ш а ю щ а я с п о с о б н о с т ь э л е к т р о ф о р е з а . Другой очень важный момент, о котором следует помнить при позиционном сопо- ставлении полос на электрофореграмме, это разрешающая способность ис- пользуемой электрофоретической системы. Оценка этого параметра имеет принципиальное значение для решения вопроса о самой возможности аде- кватного сравнения амплификационных профилей. Для того чтобы иметь такую возможность, необходимо быть уверенным, что применяемая для фракционирования амплифицированных фрагментов ДНК система позво- ляет надежно различать аллельные варианты, отличающиеся по длине как минимум на одно повторяющееся звено. Иначе можно ошибочно посчи- тать идентичными те фрагменты, которые имеют близкую, но не одинако- вую длину.
Для разделения аллелей применяемых на практике локусов с варьирую- щим числом тандемных повторов, таких как STR-локусов (длина повторя- ющейся последовательности 2—4п.н.) и VNTR-локусов с коротким шагом (в частности, минисателлитного локуса D1S80, длина повторяющейся пос- ледовательности которого составляет 16 п.н.), необходимо иметь возмож- ность различать фрагменты ДНК, которые отличаются по длине на 1—2 % (и даже меньше в случае коротких тандемных повторов). Кстати, это спра- ведливо и в случае электрофоретического фракционирования половых ге-
518
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
тероформ амелогенинового гена, при котором надежность разделения дуб- лета XY имеет принципиальное значение для использования данного теста.
Обеспечить такое разрешение способна далеко не любая электрофоре- тическая система. В первую очередь этим обусловлены высокие требова- ния, предъявляемые к электрофоретическим средам разделения.
Электрофорез в геле — стандартный метод, широко используемый для разделения и характеристики (идентификации) фрагментов ДНК в лабора- торной практике. В основе его лежит эффект «молекулярного сита», прису- щий многим гелевым средам (например, агарозным, крахмальным, агаро- вым, ПААГ). Этот феномен обеспечивает возможность функционирования молекул ДНК в электрическом поле как по величине электрического заря- да, т.е. по размеру, так и другим параметрам (например, пространственная конфигурация молекул), что в целом характеризует электрофорез в геле как сложный аналитический метод, в котором определяющую роль играет именно среда разделения. В современном анализе ДНК применяют агароз- ные и полиакриламидные гели.
Агарозные гели, безусловно, являются наиболее удобными и техноло- гичными системами для электрофоретического анализа ДНК: с ними легко манипулировать, они не обладают присущим акриламиду нейротоксичес- ким действием. Кроме того, стандартные нативные агарозные гели в гораз- до меньшей степени, чем ПААГ, чувствительны к случайным флюктуациям в макроструктуре молекул ДНК. Поэтому они менее «склонны» к опреде- ленным, связанным с этим артефактам, которые могут приводить к пози-
ционным искажениям в анализируемом геномном профиле и как следствие к ошибкам при идентификации аллелей. Однако следует помнить, что раз-
деляющая способность обычных агарозных гелей в отношении фрагментов ДНК малого размера намного ниже, чем у ПААГ. Поэтому в молекулярно-
генетических экспертных исследованиях предпочтительнее использовать специальные агарозные среды, такие как модифицированные агарозы се-
мейств NuSieveR и MetaPhorR фирмы «FMC Bio Products» (США) или ана-
логичные им продукты других фирм.
Могут применяться как нативные, так и денатурирующие акриламид- ные гели. Эти системы имеют ряд особенностей, которые должны быть предметом отдельного рассмотрения. Из принципиальных моментов отме- тим лишь то, что интерпретация электрофореграмм, полученных в денату- рирующих условиях, может осложняться эффектом «разделения цепей», когда индивидуальные фрагменты ДНК в геле оказываются представлен- ными двумя полосами. Что касается нативных ПААГ, то в целом это систе- мы с нехарактерными для агарозы электрофоретическими свойствами: они гораздо более чувствительны к пространственно-конформационным пара- метрам молекул ДНК. Поэтому в этих гелях электрофоретическое поведе- ние фрагментов ДНК значительно зависит от их нуклеотидного состава. Это может проявляться в так называемом эффекте аномальной электрофо- ретической подвижности и вызывать определенные геометрические иска- жения амплификационного профиля, в частности позиционные несоответ-
ствия, касающиеся наблюдаемых и реальных размеров фрагментов.
На практике разрешение электрофореза можно контролировать визу- ально с помощью наборов локусспецифичных аллельных маркеров (аллель- ных «лестниц») или компьютерных программных средств, используя внут- ренние или внешние маркеры молекулярных масс.
И з м е р е н и е э л е к т р о ф о р е т и ч е с к о й п о д в и ж н о с т и
ф р а г м е н т о в ДНК;
ф и з и ч е с к а я п о г р е ш н о с т ь а н а л и -
т и ч е с к о й с и с т е м ы .
Из сказанного выше очевидно, что позицион-
519
ное совпадение или несовпадение амплифицированных фрагментов ДНК в сравниваемых геномных профилях (по сути вопрос их тождественности или отличия) не всегда может быть адекватно определено «на глазок». И прежде всего потому, что при таком способе оценки трудно учесть все действую- щие факторы в их совокупности, а именно это влияет на результат. Так, на- пример, мы знаем, что при невысоком электрофоретическом разрешении можно ошибочно отождествить заведомо разные фрагменты, поскольку их позиции совпадут на электрофореграмме. Однако в более сложных случаях в сочетании с выраженным эффектом сдвига полос недостаточное разреше- ние электрофореза может привести к тому, что, наоборот, заведомо одина-
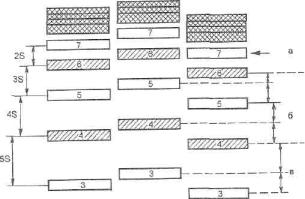
ковые фрагменты будут иметь разные позиции на геле и восприниматься как разные аллели (рис. 132).
Поэтому, строго говоря, вопрос позиционного совпадения или несовпа- дения фрагментов ДНК должен решаться аналитическим путем на том ос- новании, что позиции сравниваемых фрагментов попадают или же не попа- дают в интервалы, удовлетворяющие установленным допускам. Очевидно,
что такой подход в первую очередь требует физических измерений на электрофореграмме и учета их неизбежных неточностей.
Существует множество методов, позволяющих проводить измерение электрофоретической подвижности фрагментов ДНК на геле. При этом
вполне определенные ограничения на применение того или иного метода накладывают погрешности измерений. Так, наиболее простой, но и наиме- нее точный способ — измерение обычной линейкой. Он оказывается не- приемлемым для относительно коротких агарозных гелей, когда цена деле- ния мерной шкалы легко «вмещает» разницу в подвижности двух соседних аллелей. Наивысшие надежность и точность измерений обеспечивают авто- матические компьютеризованные сканирующие или линейные регистри- рующие системы, которые используются в наиболее совершенных прибо- рах. Более или менее универсальный вариант — это так называемый диги- тайзер (от англ. digitizer), представляющий собой электронный модуль, ко- торый состоит из курсора-измерителя и цифрового координатного устрой- ства. Такой модуль можно использовать как самостоятельно, так и в каче- стве составной части компьютерных систем обработки изображения.
Разработано много моделей подобных устройств. В качестве примера
можно привести первую в нашей стране компьютеризованную систему TVID-DATAPHOR,созданную в 1994 г. в Бюро главной судебно-медицин-ской экспертизы МЗ РФ. При помощи видеокамеры данная система позво- ляет получать и хранить в памяти компьютера «оцифрованное» изображе- ние геля. На экране монитора изображение воспроизводится с общим гео- метрическим разрешением 280 х 460 точек и монохромным тоновым разре- шением в 256 градаций. Программные средства позволяют эксперту осу- ществлять контроль электрофоретического разрешения, проводить анализ распределения материала в полосе на электрофореграмме и двумерное из- мерение пробега в геле, определять размеры фрагментов ДНК с использо- ванием нескольких алгоритмов. Предусмотрена также возможность коррек- ции электрофоретической неоднородности геля. Программа-дигитайзер имеет координатную систему с ценой деления в 1 пиксел, что в зависимости
от аппаратной оптики может обеспечить точность измерений в реальном масштабе в пределах ±0,1 мм.
Погрешности измерений — составная часть общей ошибки измерений (физическая погрешность), которая в конечном итоге определяет разре- шающую способность всей используемой аналитической системы. Кроме погрешности метода измерения, свой вклад в общую физическую ошибку
520
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
А
Б
В
I-S +S
т
Рис. 132. Решение вопроса о позиционном совпадении или несовпадении амплифи-цированных фрагментов ДНК в сравниваемых геномных профи- лях.
Этот вопрос не всегда может быть правильно решен «на глазок». Например,
при невысоком электрофоретическом разрешении в сочетании с эффектом сдвига полос можно ошибочно отождествить заведомо разные фрагменты, поскольку их позиции совпадут на электрофоре-грамме (как аллели 6 и 7 в дорожках Б и В), или наоборот, заведомо одинаковые фрагменты будут иметь разные позиции на геле и восприниматься как различные аллели (как фрагмент 5 в дорожках Б и В). Показана схематическая картина электрофо- ретического фракционирования в 3 дорожках на геле аллельной «лестницы» одного и того же полиморфного локуса с дискретным распределением алле- лей, имеющего более 8 аллельных вариантов: А — условно нормальное рас- пределение фрагментов на геле; Б — случайный сдвиг полос на величину -S (замедление); В — случайный сдвиг полос на величину +S (ускорение). В разных зонах геля в зависимости от достигнутой величины электрофоретиче- ского разрешения величина случайного сдвига (физическая погрешность системы, S) по-разномухсоотносится с величиной аллельно-го шага. Если это соотношение меньше /г, то позиционное совпадение или несовпадение фрагментов интерпретировать невозможно, поскольку это могут быть как одинаковые, так и разные аллели (зона а). Если это соотношение находится в интервале между 1Л и 1А (зона б), то можно однозначно интерпретировать только две ситуации: когда полосы полностью совпадают — в этом случае можно быть уверенным, что это идентичные аллели; когда позиции полос различаются больше чем на 2S — тогда это по определению разные аллели. Любое же несовпадение на величину, меньшую чем 2S, однозначно интер- претировать нельзя — это могут быть как одинаковые, так и разные аллели (ср., например, фрагменты 4 и 5 или 5 и 6 в дорожках Б и В). Если величина сдвига полос (S) меньше УА длины аллельного шага (зона в), то при любом
взаимном расположении полос на геле имеется возможность решить вопрос об аллельной идентичности фрагментов.
521
системы вносят такие факторы, как разрешение электрофореза, характере степень геометрических искажений в геле, форма, ширина и интенсивности самих полос и др.
Чтобы на практике оценить общую физическую погрешность исполы зуемой аналитической системы, необходимо определить так называемое окно для фрагмента, т.е. границы пространства на геле, в котором он стад тистически может оказаться. Эта величина зависит главным образом от свойств электрофоретической системы: статистического сдвига полос (т.е. величины случайных вариаций подвижности идентичных фрагментов), стен пени диффузии и разрешения электрофореза, точности измерительной сиш темы.
Отметим, что физическую погрешность можно выразить в виде доли (з процентах) от длины анализируемого фрагмента ДНК, но тогда эта величи- на будет зависеть от длины фрагмента. Например, по данным автора этой главы, на агарозных электрофоретических средах MetaPhorR в сочетаниям описанной выше системой TVID-DATAPHOR,на стандартном 12 см геле позиция фрагмента ДНК длиной 800 п.н. определяется с погрешностью! соответствующей 0,6 % длины этого фрагмента. Это означает, что такой фрагмент на разных дорожках геля попадает в «окно», аналитический разл мер которого не превышает 2 минимальных единиц измерения при достизи нутом разрешении 5 н.п. на 1 ед. измерения.
В плане решения ключевого вопроса — позиционного совпадения или несовпадения фрагментов ДНК — оценка величины физической погреш- ности используемой экспертом аналитической системы позволяет устано-1вить для данной системы соответствующие объективные правила и количе- \ ственные критерии позиционного сопоставления геномных профилей.
П о з и ц и о н н ы е к р и т е р и и с о п о с т а в л е н и я геномных п р о ф и л е й . Теоретически обобщенные требования для аналитической системы просты — позиции на геле разных аллельных фрагментов] не должны совпадать. Действительно, если бы физическая погрешность! сис- темы равнялась нулю, то полное совпадение позиций на геле двух фраг-J
ментов ДНК в противоположность любому позиционному несовпадение могло бы рассматриваться как критерий их аллельной идентичности^ Любое же несовпадение должно было бы означать разные аллели. Единст- \ венной оговоркой в этом случае будет разрешение электрофореза, которое, очевидно, должно быть способно продемонстрировать имеющуюся разницу в электрофоретической подвижности фрагментов как детектируемую (из- меряемую) геометрическую величину. Отсюда ясен физический смысл ве- <личины электрофоретического разрешения, который заключается в том, что эта величина определяет минимальную абсолютную разницу в длине] анализируемых фрагментов ДНК, которую может «показать» используемая система. Важно подчеркнуть, что эта величина зависит от длины самих iфрагментов.
Учитывая сказанное, можно было бы предложить следующие критерии.
•Два фрагмента ДНК тождественны друг другу, если их позиции на геле неразличимы при условии, что разрешение электрофореза в этой точке достаточно, чтобы зафиксировать разницу в длине между сосед- ними аллелями.
•Два фрагмента ДНК нетождественны, если при том же условии их по- зиции на геле не совпадают.
Однако не следует забывать, что физическая погрешность используемых на практике аналитических систем — реальная величина и потому позиции
522
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
идентичных фрагментов на геле вполне могут различаться — в пределах со- ответствующего «окна». Следовательно, в реальной системе два фрагмента ДНК можно считать тождественными друг другу в том случае, если, во-пер-вых, их позиции на геле не выходят за пределы границ соответствующего «окна», и, во-вторых,если заведомо разные фрагменты не могут попасть в эти границы. (Надо четко понимать, что если последнее условие не опреде- лено, то задача отождествления фрагментов на основании их позиции на геле в принципе невыполнима.)
Поскольку пространство «окна» ограничено удвоенной величиной фи- зической погрешности, то на первый взгляд можно было бы сказать так: для того чтобы иметь возможность в любом случае решить вопрос об ал- лельной идентичности фрагментов, разрешение электрофореза должно быть достаточным, чтобы величина физической погрешности не достигала Vi длины аллельного шага на минимально измеряемую величину. Однако на самом деле это справедливо только в том случае, если известны границы «окна» каждого фрагмента на геле. Поскольку эти границы априори неиз- вестны, при таком разрешении геля можно однозначно интерпретировать только две ситуации: 1) когда полосы полностью совпадают, — в этом слу- чае можно быть уверенным, что это идентичные аллели, 2) когда позиции полос различаются больше, чем на удвоенную величину физической по- грешности, — тогда это разные аллели. Любое же несовпадение на величи- ну, равную или меньшую, чем удвоенная физическая погрешность, одно- значно интерпретировать невозможно — ведь это могут быть как одинако- вые, так и разные аллели. Отметим, что это правило остается верным и в случае примененения локусспецифичных аллельных маркеров (так назы- ваемой аллельной лестницы).
Чтобы избежать подобной неопределенности и действительно при лю- бом взаимном расположении полос на геле иметь возможность решить во- прос об аллельной идентичности фрагментов, разрешение электрофореза должно быть таким, чтобы величина физической погрешности системы оказалась меньше 1А длины аллельного шага на минимально измеряемую величину.
Таким образом, мы сформулировали универсальные критерии.
АДва фрагмента ДНК тождественны друг другу, если их позиции на геле не отличаются более чем на удвоенную величину физической по- грешности системы при условии, что разрешение электрофореза в этой точке достаточно, чтобы величина физической погрешности
была меньше У\ длины аллельного шага на минимально измеряемую величину;
▲ Два фрагмента ДНК нетождественны, если при том же условии их по- зиции на геле отличаются более чем на удвоенную величину физичес- кой погрешности.
С р а в н е н и е г е н о м н ы х п р о ф и л е й , п о л у ч е н н ы х на р а з н ы х г е ля х . По существу сопоставление геномных профилей, по-
лученных на разных гелях, означает, что сравнению подвергаются результа- ты разных экспериментов. Этот факт имеет принципиальное значение: если при сравнении позиций фрагментов ДНК на одной электрофореграм-
ме их позиционное совпадение или несовпадение устанавливают путем прямого (непосредственного) сопоставления друг с другом, то при анализе разных электрофореграмм этого сделать нельзя, поскольку профили элек- трофоретического разделения в каждом эксперименте оказываются не- одинаковыми. На разных гелях амплификационные профили идентичных
523
объектов могут быть весьма похожими, но они необязательно будут геомет-'рически тождественны. Поэтому сравнение можно делать только опосредо- ванно — путем независимого сопоставления параметров сравниваемых полос с некой третьей величиной. Далее известно, если две величины по- рознь равны третьей, они равны между собой и т.д.
Совершенно очевидно, что таким согласующим параметром является
аллельная принадлежность фрагментов в сравниваемых геномных профи-
лях. Сопоставление аллелей (генотипов) в отличие от позиционного срав- нения амплификационных профилей имеет уже не относительный, а аб- солютный характер, поэтому генотипированные аллельные профили в принципе можно сравнивать независимо от того, получены они в одном или в разных экспериментах. Это означает, что генотипические характе- ристики исследованных объектов могут быть организованы в информаци-
онную базу данных и использоваться как элементы банка генетических данных.
Для индивидуализирующих систем, основанных на использовании по- лиморфных локусов с дискретным распределением аллелей, применение локусспецифичных аллельных маркеров (аллельных «лестниц») позволяет
относительно просто осуществлять генотипирование локальных аллельных профилей и при условии, что в каждом эксперименте были соблюдены не- обходимые позиционные критерии сопоставления полос (см. выше), пере- ходить от позиционного анализа в рамках одного эксперимента к безотно- сительному сравнению генотипов объектов.
В том случае, если используются другие (неаллельные) маркеры молеку- лярных масс, первичной согласующей величиной для сравнения на разных гелях амплифицированных фрагментов ДНК является длина этих фрагмен- тов. Однако в этом случае понадобятся дополнительные условия, которые обсуждаются в следующем разделе.
О п р е д е л е н и е р а з м е р а ф р а г м е н т о в и г е н о т и п и р о - в а н и е г е н о м н ы х п р о ф и л е й . Физические измерения пробега
фрагментов ДНК на электрофореграмме дают возможность вычислять их условный молекулярный размер, например длину в парах нуклеотидов. Между этими двумя параметрами существует вполне определенная взаимо- связь: молекулярная масса (а следовательно, и размер) обратно пропорцио- нальна величине электрофоретической подвижности. Надо подчеркнуть, что эта зависимость имеет сложный характер, она не является линейной и
ее конкретные характеристики могут существенно различаться на разных гелях. Последнее объсняется тем, что электрофоретическая подвижность молекул нуклеиновых кислот в геле, в частности ДНК, определяется целым рядом параметров, таких как молекулярная масса, линейный размер моле- кул, а также молекулярные структурные особенности высших порядков. Вклад каждого из факторов и, следовательно, форма кривой, характеризую- щей эту многопараметрическую зависимость, могут быть разными: все за- висит от аналитических свойств конкретной гель-электрофорезной сис-
темы
Таким образом, чтобы определить размеры амплифицированных фраг- ментов ДНК по их расположению на геле, нужно проанализировать про-
филь электрофоретического разделения для данного конкретного геля и для него определить точный характер зависимости электрофоретической подвижности фрагментов от их размеров. Это можно сделать, используя стандарты молекулярных масс, т.е. фрагменты ДНК заранее известного размера, которые фракционируют на том же геле, что и анализируемые ам- плифицированные фрагменты.
524
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/