

3 курс / Патологическая физиология / Основы общей патологии
.pdf! |
681 |
Передобсуждениемпр облемаутоиммунныхзаб, леванийтметим, чтоаутореактивныеВ -лимфоцитынепроходятподобногоэтапа негативнойтимичсел. скойкции
Идентинелимфоидныхикацияклеток
Клеткимогутбытьидентифицирпоморфолованыгическим |
|
|
|||
функцриональнымтер |
|
иям.Моноклональнаятехнология |
|
||
полученантитпроиеволюциюязэтела,мтакпросекак |
|
|
|
||
теперьмногиеклеткимогутбытьопозразделеныа(иногда)по |
|
|
[398] против |
||
молекуламклеточнойповерхностиаркерам(), |
моноклональныеант |
||||
котмополучитьрыхжно |
|
|
итела (X. Кёлер, |
||
С. Мильстейн, Терм1983)маркер»«спон. дльзуется |
|
|
|||
обознклеточныхачениянтигенов,котоспособныреагироватьые |
|
|
|||
соспецифант.Вичтескимилами |
|
таблице 24,приведённойнас. |
|||
400-401,представленычащевсегоиспользуемыемаркеры,не |
|
|
|||
толькодляТ |
- иВ |
-клеток,ноидляэффекиммунногоответа:оров |
|
|
|
моноцит,гранулоцикровяных, пласестественныхинок |
|
|
|||
киллеровК(N |
-клетки);последниеиграютважнуюроль |
|
|
||
противоопухолевомммуните |
тесм.раздел( Воспаление«»). |
|
|||
СокCDращениеекомендованоВОЗпроисходитотанглийского |
|
{16}.Знакомясь |
|||
«Cluster designation» — «Обозначениескопления» |
|
||||
сэтойаблицей,читавстнретитлькоторыеобозначени |
|
|
ямолекул |
||
клеточнадгезии,знакомыепопредыдущимйтаблицанас. |
|
|
14-16). |
||
Иммуноглобулиныкакмараспознающиекерымолекулы |
|
|
|||
Иммуноглобулины (Ig)или,чтожесамое,антитела, |
|
-клетокили |
|||
являютсяпродфуктаминкциональноактивныхВ |
тоятизполипептидныхцепейразличной |
|
|||
плазмацитов.Онисос |
|
|
|
|
|
конфигурации.Имеетсяосновныхклассов5 или, ( |
|
|
|
|
|
иммунологическихизотипов) разлIg,такжеподклассычают. |
|
|
|
||
Существованиеантител,какоторвавш« кл ткихся |
Эрлих(1901)Структура. |
||||
рецепторов»постулировалвпервыеП. |
|
|
|||
иммунбыла,воглснчео,булиноврасшифрованавныхтах |
-1962). |
||||
Р. Р. ПортеромиДж. |
М. |
Эйдельманом(1957 |
|||
МолекулысостоятIgизполипептидныхцепей,соединенных
! |
Каждаятяжилецепьгкаялаяявляетсяполипептидом, |
683 |
|
||
сформированпутемтранскрипцииопределенногосляцымй |
|
|
|||
количествагенов,кодирующихпостоянныевариабельныеучастки |
генногосплайсинга |
|
|||
Ig,посредствомсложнойсисте ы |
|
||||
(С. Тонегава, 1991)Легкиецеписодержат.каквари, ибельныек |
|
||||
констуч,первасннаходятсякиыNHе |
2- концемолекулы. |
||||
Сущлецепигкиествуютдвухклассов:κ |
- иλ -.Тяжелыецепитакже |
|
|||
содеваржучасткииабельныевNH |
2-концахконстучаснткиные |
|
|||
наCOOH -концах.Существуетклассили5 изотиповтяжелых |
|
||||
цепей,аименно:γ,α,μ,δ,ε |
|
|
— соответственнодляIgG, IgA, IgM, |
|
|
IgDиIgE. |
|
|
|
|
|
|
РазницамеждуконстчасНнтныками |
-цепейобъясняет |
|||
различныебиологичефункцииэтихклассовIgкие. |
|
|
|||
|
• IgG – наиболееширокора |
спространмономернкласс ый |
|
||
иммуногсыворо,элобулектмкрофоигрирующийнв етически |
|
|
|||
вееγ |
|
|
-фракции.Этоважныйпротивовирусный |
|
|
противобактерфакторпотенцопсонин,атакжеиальный |
|
|
|||
главныйучастнвториммуннкответачногоединственныйго |
|
|
|||
классантител |
|
- нейтралокс.ЭтоуникальныйзаторновIg, |
|
||
проходящийчерезплацентуспособныйобеспечпассивныйать |
|
|
|||
иммуниноворожденнымотдо3етмесяцев6 .Егоприблизительный |
— 3недели,онявляетсяоднимиз |
|
|||
периодполураспадавсыворотке |
|
||||
двухклас |
совIgкоторые.активируюткомпполассическомуемент |
4.Они |
|||
пути.Существуетподкласса4 IgG: IgG |
1, IgG2, IgG3,иIgG |
||||
отличаютсянебольшидеталяН ми |
-цепиFc -фрагмента.Антитела |
|
|||
IgG2 чащеявляютсяопсионинамитвечаютпопаданиетоксинов |
|
1 и |
|||
идекстранов. |
Анти-Rh-антителачащеявляютсяподклассамиIgG |
||||
IgG2. IgG4 учеловекаимеютзначительнропизмктучнымй |
|
||||
клеткам,втовре,какIgGя |
|
1 неспосвязыватьсябны |
— дело |
||
мастоцита,угрызун,вчастно,мсвинкиврскойти |
|
||||
обстна.оборотит |
|
|
|
||
|
• IgAсм.т |
( |
акжевыше,с. |
386)учеловекасыворотке |
|
присутствуетнакак80%мономер.Умлекопитающихэто,в |
|
|
|||
основном,димер. |
|
[400] [401] [402] УIgAимеютсяспецифические |
главный |
||
черты,связанныесослизистымиоболочками. IgA |
— |
||||
секреторныйпродуктсистемылимфоидной |
тканиверхнихнижних |
|
|||
! |
685 |
|
В-лимфоцитов(1994). |
|
|
• IgE — этореаилгомоцитотропныеи антитела.Кклассу |
|
|
IgEотносятсяклассическиекожно |
-тропныеантитела,играющие |
[403] |
важнуюрольвреакцияхнафилактической |
||
гиперчувствитетипаI.Большинствоихбио огическихьности |
|
|
свойствсвязанотемфактом,чтоFc |
-фрагцепиэтойментолекулы |
|
оченьпрочносвязываетсяεFc |
-Iрецепторамитучныхклеток |
|
(мастоцитов)базофилов.Рец |
епторыIgEболеенизкср,одстваго |
|
походуанафил,возникаютакуэозжесиинофилов |
|
|
тромбоцитов.КогдаFab |
-фрагментэтих,связанныхклеткойIgE |
|
специфическиперекрысоотванается,ствующимигеном |
торов |
|
происходитдегрануляцияклетоквысвобождениемедиа |
||
анафилакснебольших.В количествIgEприсутствуетплахзме |
|
|
кровитканях.Налимфоцитахмонаходитсядругой |
|
|
рецепторкIgE (FcR |
-2),образующийгораздоменеепрочнуюсвязь. |
|
В таблице 25 приведеныосновныесвойствафункцииклассов |
|
|
подклассовIg. |
|
|
Разнообантителвыравтом,чтозиежкаетсяждый |
|
|
индивидуальныйВ |
-лимфвсеегоклональныецитпотомки |
- |
вырабатываютиммуноглобулинсуникальнойструктуройантиген |
||
связывающихучастков, |
тоестьидиотипынеповторяются. |
|
ВариабельныеучасткикодируютсяV |
-генами. |
|
Этобылоневобеспечитьзможно,еслибыкаждый |
|
|
иммуноглкодиротначала,доконцабувасвоеймолекулыинся |
|
|
отдельнымгеном.С. |
Тонегава(1988)установил,чтогетерогенность |
|
антителобеспечивблагодарятому,чтоотдельныеетсоставныея |
|
|
частикаждизполипептидовхгокодируютсяразныгенами |
|
|
зародышевлин,любойикотисуществуеторыхйкак |
|
|
множественныймультигенподвергаетсясоматическому |
|
|
мутирован.Большоезначениепр ю |
возникразновенииобразия |
|
антителтакже( Т |
-клеточныхрецепторов)имеюттранспозоны |
— |
мобдильныесперучасткигенир,способныеомваенять |
|
|
местопвхроБмл.(ожениесомах |
МакКлинт, 1983)Оборотной. к |
арьирования |
сторонойработыэтогоэффективногомеханизма |
||
программявляетотносивысокийярисктаельтранспозицийих, |
|
|
которыеприводятэкспрессиилимфоклетнк.дныхогеновах |
|
|
! |
• Дляк |
онстантныхрегионовтяжелыхцепейтакжеиме тся |
687 |
|
|
|
|||
нескольковариантальтернагенот, квотзависиторыхивных |
|
|
|
|
|
||||
принадлежностьантителодантигеннойспецифичностикразным |
|
|
-клетокможет |
|
|||||
подклассами .ОдинтотжеклонВ |
|
[404] со |
|
|
|
||||
переключатьсинтез |
|
дногоклассаиподклантителсса |
|
|
|||||
другой,неменяяспецифичности,путёмполиаденилирования |
|
|
-копииразныхгенов, |
|
|
||||
первичноготранскрипта,содержащегоРНК |
|
|
|
|
|
||||
альтернативныхточках,отчеготранскриптприсплайсингетеряет |
|
|
|
|
|
||||
разныеучастки. |
|
|
|
|
|
|
|
|
|
|
БиологияТ |
-лимфоцитов |
|
|
|
|
|
||
|
Тимус-завислимфоцитыилимыеТ |
|
|
-клеткиявляются |
|
|
|||
основнымикомпонениммун.сОтнеаойимыи |
|
|
|
|
|
|
|||
иммуноспецифичнысобныбеспечиватьиммунологическую |
|
|
|
|
|
||||
памятьифункционироватьнесколькихрегуляторных |
|
|
|
яв |
|
||||
эффекторныхмоделях.Основнойпредпосылкойиучасти |
|
|
|
- |
|||||
иммунномответеслужитТ |
|
-клеточраспознаваниеантигенаое.Т |
|
||||||
клеткиональнограниченырестриктированы( ),поскольку |
|
|
|
|
|
||||
каждаяизнихсодержитуникальныйрецепт,спосорбный |
|
|
|
|
- |
||||
взаимодействоватьопределенантиге.У Т95%нымом |
|
|
тор (TcR)состоитизα |
- иβ |
|||||
лимфоцитов Т-клеточныйрецеп |
|
- |
|||||||
полипептидныхцепей,сконстучас,расположеннымиками |
|
|
|
|
|
||||
ближекповклеткирхности,вар ,удаленнымиабельнымиот |
|
|
|
|
|
||||
поверхнклетки,котсостиедрыеуникальнымняются |
|
|
|
делова |
- |
||||
антиген.Благразличовструкдарямиюотуреальных |
|
|
|
||||||
иβ -цепей,т..полиморфизмувсемействеTcR,возможноразвитие |
Дэвис, 1988)Механизмыгенерации. |
|
|
||||||
различныхклоновТ |
|
|
-клетокМ(. |
|
|
||||
этогоразнообразияаналогичописаннымвышедля |
|
|
|
|
|
|
|||
иммун,стразницойглобулинов,чтопергенетическихйтасовка |
|
|
|
|
|
||||
компонентов,кодирующихразличныеэлементыTcR,вовлекает |
|
|
|
|
|
||||
хромосомыи7Целая14мол. црецепторапикулаимеет |
|
|
|
|
|
||||
трансмембранныйучастокицитоплазматическийхвост.Последний |
|
|
|
|
|
||||
исподпередачильзуетсясигвнклеткиутрьала.Вцеломэта |
|
труктурусвязанногоклеткойиIg, |
|
|
|||||
структураочепохожань |
|
|
|
|
|
||||
TcR,такжекакимолекулы1 |
|
|
-гои2 -гоклассаГКГС,являются |
|
|
||||
членамисуперсемействаIg |
-генов( |
рис. 86). |
|
|
|||||
|
Недавно,былаидентифицированачастьТ |
|
|
-клеток,которые. |
|
||||
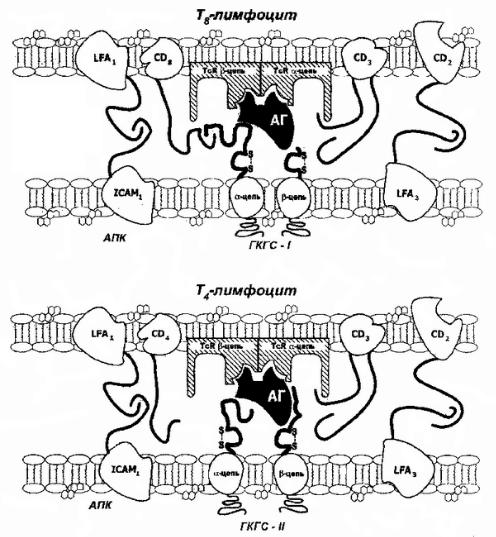
! |
689 |
Рис. 86Взаимодействие. Т |
-клетоксанти |
геномнаантиген |
-презентирующихклеткахАПК(). |
Объяснениявтексте. |
|
|
|
Т-клеткиразличаютсяпофункцимаркеры,различныих |
таблице 24.Втимусе |
|
|||
клеточнповерхности,каквышейказано |
|
|
|||
незрелые Т-клеткиэкспрессируютТ |
-клеточно-специфичные |
||||
поверхностныеантакие, игеныкакОниCD2образуют. |
|
|
|
||
вспомо,лекувчастностигате, CD4лCD8ьные. |
|
|
|
|
|
КогдазрелыеТ |
-клетпокти,онидаютмусявляютсяибо |
-положительными.Т |
- |
||
СD2/СD4 |
-положительны-ми,либоСD2/СD8 |
||||
клеткиразвразличниваютсяклоны,каждыйизкоторыхнесет |
|
|
|
||
отличнуюдругихмолекулуспецифичнTcR,особомую |
-1илиТII. |
-клетки, |
|||
комплексуантигенагликопротеидаГКГС |
[405] |
||||
неспособныереагировать |
собственнымиГКГС |
|
- |
||
гликопротеидамибесполе( |
зные)уничтожаютсяотнедостаточной( |
|
|
||