

3 курс / Патологическая физиология / Основы общей патологии
.pdf! |
|
|
671 |
|
|
|
Клеткамипамятиназываютвселимфоциты,котп явилисьрые |
|
|
|
|
||
послепервичногоиммунногоотв, тазультклональнойте |
|
|
|
|
||
экспансии,индуцировантиге.Прикосантактеомнойтигном |
|
внениюсродоначальником, |
|
|||
ониспособныкусиленному, сра |
|
|
|
|||
отвенаан.ЭтоуиТ,иВген |
|
|
|
-клетки.В |
-лимфоцитыпри |
|
вторичиммунномответеачинаютантитело |
|
|
|
-образованиенес |
||
иммуноглобулиновМ,какихпредки,асразуиммуноглобулинов |
|
|
|
|
||
G. |
|
|
|
|
|
|
Таккакбольшантнесшиваетнствогеновнеп сре |
|
|
клеткиспособнытветить |
дственно |
||
иммуноглобулиновыерецепторы, Т |
|
|
|
|||
лишьнасеквенциальдетермиант,предснгеаыенатавленногоы |
|
главногокомплекса |
|
|||
АПКвассоцгликопротеидамиации |
|
|
- |
|||
гистосовместимости (ГКГС) |
- ГКГС-антиIклассовIIгенами.В |
|||||
клеткира |
спознаютконформацдетерминтактоннаныетыого |
|
|
|
||
антиге,впри,могутнсамостоятельноципеотвечанаэпи, опы |
|
|
|
|
||
котомплерымихиммуноглентарнырецепторы,нобулиновые |
|
|
|
- |
||
дляполноценного,масштабногоответануждаютсяпомТ щи |
|
|
|
|
||
клеток. |
|
|
|
|
|
|
Антиген-презентирующиефункциивыполняет,основном, |
|
|
|
|||
системамононуклеарныхфагоцидругиев дытовток |
|
|
|
|
|
|
(эндотелиальные,гли),кртогозвездчатыемеклетки |
|
|
|
|
|
|
корытимусаэктодермального( происхождения)Основными. |
|
|
-лимфоцитов |
|||
клетками,процессантдляиВрующимигены |
|
|
|
|||
считаются |
дендритныеретикулярныеклетки |
|
лимфоидных |
|||
фолликулов,адляТ |
-лимфоцитов |
— интердигитирующие |
||||
ретикулярныеклетки |
паракзлимфоузловртикальных. |
|
|
|||
КлоныАПКнеспецинобрализированыкакб ткуго |
|
|
|
-то |
||
одногоантигена.Онимогутпредставлятьлю |
|
|
|
быеуникальные |
||
антигеныэкзогеннэндогенного( пр исхождения,чужеродные |
|
-клеткамсоответствующей |
||||
аутоантигены) |
-отвечающимТ |
|||||
клональнойспецифичности.В |
|
-клеткатакжеспособнапредставлять |
|
ако,в |
||
антигенвассоциацииГКГС |
|
-IиГКГС |
-IIмолекулами,одн |
|||
отличие |
[391] |
АПК,онаклонально«рестриктирована» |
-лимфоцит |
|||
определенИнымиантиге. словамин,каждыйомВ |
|
|
|
|||
можпредвассоциацииттавлятьГКГС |
|
|
|
-IиГКГС |
-II |
|
гликопротевобработанномвиделишьтотуникальныйдами |
|
|
|
|
||
! |
673 |
|
|
|
|
|
|
|
работают,к утокипаракринныерегулят.Ввысокихры |
|
|
|
|
|
|
|
|
дозах,образуясьприполиактивлональнойции |
|
|
|
|
|
|
|
|
иммунокомпетентныхклеток,некотизнихрыегутказывать |
|
|
|
|
|
|
||
системэффект.Униый |
|
кальнцито,какпаракринныхстьинов |
|
|
|
- |
||
регуля,заключаетсявто,чтрбмольшинствоихклеток |
|
|
|
|
|
|
||
мишенейциркули.Поэтчерезоуюткальныемузоныпродукции |
|
|
|
|
|
|
||
цитокиновмогутпроход,активситьстемнороваться |
|
|
|
|
|
|
|
|
распрбоколичествальшиестранятьсякл. ток |
|
|
|
|
|
|
|
|
Основныеизцитокиновохарактеризованытаблице( |
|
|
|
Табл. |
23, |
|||
см.такжеглаву |
«Патофизиологиявоспаления» |
|
|
|
)Они. |
|
|
|
вырабатываютсямножкл,спосествомк |
|
|
обныактивировать |
|
|
|
||
продукциюдруга,покаскадпри,имеютнширокийципуому |
|
|
|
-1,ИЛ -6и |
||||
кругчастичноперекрывающихсяфункций.Например,ИЛ |
|
|
-2иИЛ |
|||||
ИЛ-8являютсяпровоспалительными;ИЛ |
-9 — факторы, |
|||||||
стимулирующиеростлимфоцитов;ИЛ |
|
-4иИЛ |
-5вовлекаются |
|
|
|||
переключениесинтезаклассовIg;ИЛ |
|
-10инте,чторесенонм |
|
|
||||
можсинтезпрекращцито, акиобразомтьобеспечиваянов |
|
|
|
|
|
|
||
отрицателобратнуюсвязь.Вразделе,посвященномнуюответу |
|
|
|
|
- |
|||
остройфазы,мыужекасаливойнекоторыхсцитокиновьтвИЛ( |
|
|
|
|
|
|
||
1,ИЛ -6,ИЛ -8,ФИОидр.)Интегра. резуработыльтатомным |
|
|
|
|
|
|
||
системыцитокиновявляетсяорганизацияпре ммунного |
|
|
|
|
|
|
||
иммунногоотв,регуляциятаихс цитотоксическоголы |
|
|
|
|
|
|
||
потенциала,возможно |
|
— итерм.Циигрнацток, аикютяны |
уюрольвхроническом |
|
|
|||
показановпредыразгигантскдущихелах |
|
|
|
|
||||
воспал,генлихорадкиезениирегуляциигемостаза,обладают |
|
|
|
|
|
|
|
|
гормонопэффеки о,какдгутэтообаминымисуждаетсяниже, |
|
|
|
|
|
|
||
участвоватьразвитиистресса. |
|
|
|
|
|
|
|
|
• Интерфероны |
(ИФН)являютсягруппойразнородных |
|
|
|
|
|
|
|
цитокинов,которыебыливыд |
|
леизнадосаыжидкостиочной |
|
|
|
|
|
|
культурклеток,инфицированныхвирусами.Этиж ,дкости |
|
|
|
|
|
|
|
|
смешанныеклетками,инфицированнымидругимвирусом, |
|
|
|
|
|
|
||
тормозилиразмножепослед.ИФНпредставляютниеего |
|
|
|
|
|
|
||
естественныепротивовбелкинтересн,каксигналыру, е |
|
|
|
|
|
|
|
|
включающиезащитнуюпрограммуприв русномповреждении |
|
|
|
|
|
|
||
клетки.Посамкольку |
[392] [393] [394] вирycпредставляетсобой |
|
|
|||||
патогеннуюпрограмму,интерферзащитаос ованановая |
|
|
|
|
|
|
|
|
торможенииеёсчитываниякопирования. |
|
|
|
|
|
|
|
|
! |
675 |
чтоусиленнобразэкзогенноевведениеваниеслечц бнойлью различныхИФНобладаетотчётливымпровоцирующимдействием отношенииаутоиммунныхзаб, левасобе,тироидитаийно Хашимотодругихэндокринопатий(1983).
ИΦΗγобладаменеевыраженнымпротивовирусным |
|
|
|
||
действием.Ноиз |
-засильногоиндуцирующегоэффектана |
сию |
|||
цитотокспозначительнособичусиливатьностэкспрес |
|
|
|||
антигеновГКГС,кактакI, 11ласса,именноэтотИФН |
|
|
|
|
|
представнаибольшийиммунологическийяетинтерес.Этот |
|
|
|
||
медиатмалогомологдрИФНугим,имерецепторчнойи |
|
|
|
|
|
кодируетсядругхр мосомойдевят()Он.пр изводится |
мисяТ |
-клетками,втомчислеТ |
- |
||
активированнымипокоящи |
|||||
хелпертип1,атакженормамикиявляется, лерамиьны |
|
|
-4Он. |
|
|
главнымктивамакрофагов,такжеантагонистомромИЛ |
|
|
|
||
стимоул2екирулыет |
|
-гоклассаГКГСковзаимодействиюсо |
|
||
многклеткамиможетбытьсинергистомфакт |
|
|
оранекроза |
|
|
опухолиФНО)ИФНγ(. считаетсяусилителемклеточного |
|
|
|
|
|
аутоиммунитета,вжевремяон мозитяд |
|
|
|
|
|
проявлеиммуг нимораль,вчастностийтета,синтезIgE,вого |
|
|
|
|
|
большихдозах |
— иантителдругихклассов.Счи, тоается |
|
|
|
|
наследственныйпосттрансдефектвсекрИФНγипцециионный |
|
|
|
||
— важнпреанафийшаядпосыреактивностилактическойка. |
|
|
|
||
ТаккакрецепторИΦΗαβкодируетх ом21,б сльныеома |
|
|
|
|
|
трисомиейподаннхромосомесиндром( Дауна)располагают |
|
|
ительностьюкэтим |
|
|
избыткомрецепторовотличаютсячувств |
|
|
|
|
|
цитокинам,чтовызываантипролиферативноеусиленное |
|
|
|
||
иммуносупрессорноедействиеИФН,манифестирующееввиде |
|
тимуса,аутоиммунных |
|
||
лимфоп,раннатрофейниии |
|
[395] |
|
||
синдромов,повышчастотылейкоза,компенсаторногонн |
|
|
|
||
сниженияуровня |
ИФНвкрови |
|
|
|
|
• Множество колониестимулирующихфакторов |
(КСФ),таких |
|
|||
какгранулоцитарныйгранулоцитарно |
|
|
-моноцитарный,также |
|
|
производятсяТ |
-клеткамиЭтифакторыобеспечивают |
|
|
||
взаимеждулимфоиднойодействиегемопоэтическойсистемами, |
|
|
|
||
участвуютпатогенезе |
|
лейкоцисдвядерниформулытг взойв |
|
|
|
гранулоцитов,характерныхдлявоспалении |
|
|
|
|
|


! |
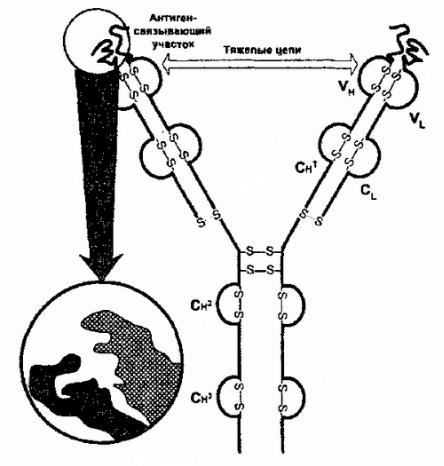
Молекулы 1-гоклассаГКГС |
677 |
(рис. 83) — этосвязанные |
||||
мембранойгликопротеиды |
|
|
|
|
|||
представленнабольшиядроныестве |
-цепикоторая |
||||||
содержащихклет.Онисостояткбольшойз |
|
|
|
|
|||
соединенанековалентнойязьюβ, |
|
|
|
-микроглобулиномα |
-цепь — |
||
высокополиморфна,еевариабельныеучасткинаибоудаленые |
|
|
|
|
тичбезски |
||
отклеточноймембраныМономорфныепрак( |
-цепиβ |
|
|
|
|||
полиморфизма)частиα |
2-микpoглбyлинaприближенык |
|
|||||
клеточноймембране |
|
|
|
|
|
|
|
Рис. 83Строение. белковглавногокомплексагистосовмеIIIкласса. тимости
Данныемолекулыучаствуютраспознаванииантигенов, |
|
|
D8-положительными |
||||
синтезируемыхвнутриклеток, C |
|
|
|
|
|||
цитотоксическилимфоцитаАнтиген,вырабатывающийсями |
|
|
|
|
|
||
клетке — безразлично,свойилипаразитарный,напримервирусный |
|
|
|
|
— |
||
— процессируетсяособымкомпнелексизосомпротеазльных |
|
|
|
|
|||
протеасомой.Вшерохэндоватомплазретикулюмеатическом |
|
|
|||||
(ШЭР)синтезируютсябелкиГКГСкласзатемI , ониоединяются |
|
|
|
— |
своих) |
||
сфрагментами |
эндоцеллюлярных (необязательно |
||||||
антигенов,поступающихизпротеасомВэтомучаствуют |
— ABC-белки.КомплекспептидаГКГС |
|
|
||||
специальныепереносчики |
|
|
|
||||
Iивнутриклеточногоанти |
генапоступаетваппаратГольджи, |
|
8- |
||||
оттуда — наружнуюмембрануАПК,дляпредставленияСD |
[396] |
|
|||||
положительнымцитотоксическимлимфоцитам. |
|
|
|
||||
Молекулыкласса2 |
|
-гоГКГС |
(рис. 83)содержатковалентно |
|
|||
соединенныеαиβ |
|
|
|
|
|
|
|
|
-цепи,являющиесявысокополиморфными |
|
|
|
|||
белками,похожиминаα |
|
-цепикласса1Устройство. полиморфных |
|
|
|||
мономорфныхучастковблизкоописанномуклассаГКГС1. |
|
|
|
|
|
|
|
Этимолекулы |
|
— непременныеучастникираспознавания |
|
|
|||
! |
679 |
Описанныесобытиясоставляютсутьпроцессингаантигенов
антигенпредставляющихклетках. |
|
||
|
Всетипа4молекулклассаи1ГКГС2,вмесант,Тителами |
- |
|
клеточнымиантигеннымирецепторами, Fc |
-рецепан,тителорами |
||
CD2-4 иСD 8-рецепторамилимфоци,секреторнымфрагментомв |
|
||
IgA — являютсячленаминадсг ммуноглобулиновмейства, |
|
||
ихконфигурациянедалекаоттипичмолекулыанойтитела |
ихпредшественников |
||
(рис. 83)Всеэти. молекулыпроизошлиотобщ |
|||
|
|
|
|
иимеютмножеспептпетлидисульфидныеве,связи |
|
||
структуре.Молекулыкласса3 негомэтнадсемействуологичныму. |
|
||
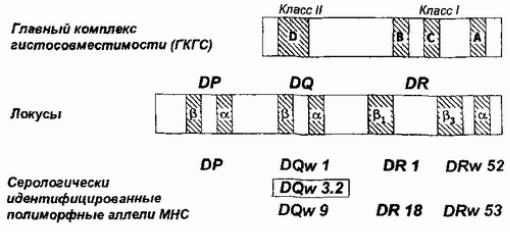
ВышеужеговосцеплегенамирилосьмеждуГКГС |
Рисунок 84 |
||
нек,остаутоиммубеннорыми, лез. нямиными |
|||
иллюстрируетподобнуюсвязьнапр нсулинмере |
-зависимого |
||
сахарногодиабетатипа1. |
|
||
Рис. 84Геныглавного. комплексагистосгеныопределяющвм стимостие подверженностьинсулин -зависимомудиабетуI ипа. ВариаDQ,тессцепленныйотдиабетомуказанврамке.
Теорияклональнойселекцпроиразвитиясхождения иммунныхклеток
Этатеопроияразвитиясхождеиммуклетокбыланныхия
сформулирв1957годунезависимодругдругатванаМ. |
Бернетом |
вАв стралииД. |
ТолмейджемвСША.Частьизеепостулатов |