

25. Условия возникновения оптимума и пессимума частоты и силы раздражения (н.Е. Введенский).
При сравнительно низких частотах раздражения наступает зубчатый тетанус, при большой частоте – гладкий тетанус. При зубчатом тетанусе каждый последующий нервный импульс воздействует на начавшую расслабляться мышцу, при этом происходит неполная суммация сокращений. При гладком тетанусе, имеющем бо́льшую амплитуду, воздействие импульса происходит в конце периода укорочения, что приводит к полной суммации сокращений.
Если ритм раздражения увеличить примерно до 10 - 15 раз в 1 сек, следующие друг за другом по нерву раздражающие сигналы сближаются во времени настолько, что каждый последующий стимул будет воздействовать на мышцу до того, как она успеет полностью расслабиться после предшествующего сокращения. Тогда в ответ на второе, третье и последующие раздражения мышца будет сокращаться, исходя не из полностью, а только частично расслабленного состояния. Это позволит записать неполный, или зубчатый, тетанус.
П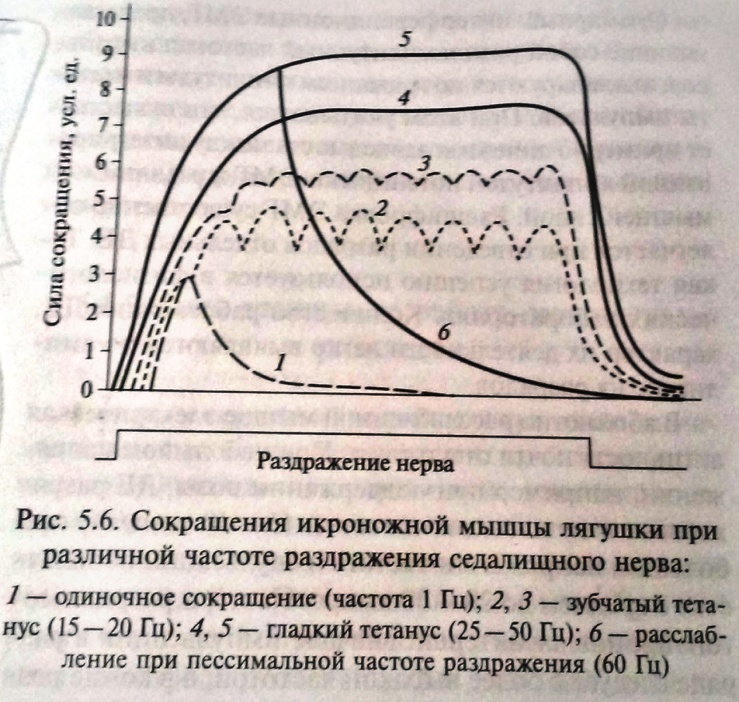 ри
еще большем ритме раздражения (для
икроножной мышцы лягушки не меньше 20
раз в 1 сек) следующие друг за другом
импульсы сближаются в такой мере, что
каждый последующий действует раньше,
чем закончится восходящая часть
предыдущего сокращения. И поскольку
интервал между стимулами короче, чем
фаза сокращения, расслабления в интервале
между раздражениями вообще не происходит.
Результатом является сплошной
(гладкий) тетанус,
характеризуемый длительным сокращением,
не прерываемым расслаблениями.
ри
еще большем ритме раздражения (для
икроножной мышцы лягушки не меньше 20
раз в 1 сек) следующие друг за другом
импульсы сближаются в такой мере, что
каждый последующий действует раньше,
чем закончится восходящая часть
предыдущего сокращения. И поскольку
интервал между стимулами короче, чем
фаза сокращения, расслабления в интервале
между раздражениями вообще не происходит.
Результатом является сплошной
(гладкий) тетанус,
характеризуемый длительным сокращением,
не прерываемым расслаблениями.
Оптимум и пессимум частоты раздражения. Н. Е. Введенский показал, что величина тетануса может значительно отличаться от величины одиночных сокращений, в зависимости от силы и частоты импульсов. Умеренные по силе и частоте раздражения (оптимальные), вызывают максимальный эффект, значительно превышающий амплитуду одиночного сокращения, а очень сильные и частые – (пессимальные), вызывают заметное ослабление эффекта.
На основании этого Н. Е. Введенский сделал вывод, что высота тетануса определяется не только наложением отдельных сокращений друг на друга, но и теми функциональными изменениями, которые оставляют в ткани приходящие раздражения. Если каждый последующий импульс приходит с таким интервалом, что застает ткань в состоянии повышенной реактивной способности, то эффект сокращения будет сильным, а высота тетануса больше. Н. Е. Введенский назвал это состояние повышенной возбудимости вслед за протекшим сократительным эффектом экзальтационной фазой. Экзальтационной фазе предшествует состояние пониженной реактивной способности – рефрактерная фаза. Импульсы, следующие с такой частотой, при которой они попадают в рефрактерную фазу, вызывают пессимальный эффект, снижение высоты тетануса. Это явление называют пессимальным торможением.
Таким образом, пессимальным или оптимальным будет ответ, зависит от того, в каком состоянии находится ткань к моменту прихода действующего на нее импульса. Для свежей, неутомленной икроножной мышцы лягушки оптимальная частота раздражений, дающая при физиологически максимальной силе раздражения тетанус наибольшей величины, составляет 50-100 раз в1 сек; увеличение частоты раздражения до 200 - 300 раз в 1 сек приводит к пессимальному эффекту.