

2 курс / Нормальная физиология / Физиология_дыхания_Частоедова_И_А_,_Еликов_А_В_
.pdf41
Скорость доставки кислорода в нормальных условиях значительно превышает его потребление, в результате чего лишь малая доля доступного кислорода извлекается из капиллярной крови в обычном состоянии (в покое КУО2 = 25-35%). Это позволяет тканям приспосабливаться к снижению доставки кислорода
Зависимость степени оксигенации гемоглобина от парциального давления О2
в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина, или сатурационной кривой (рис. 15).
Рис. 15. Кривая диссоциации оксигемоглобина, или сатурационная кривая
Плато кривой диссоциации характерно для насыщенной О2
(сатурированной) артериальной крови, а крутая нисходящая часть кривой — венозной, или десатурированной, крови в тканях. Кривая имеет S-образную форму, что обусловлено кооперативным взаимодействием четырех субъединиц,
составляющих тетрамер гемоглобина. Присоединение О2 к гемму одной субъединицы повышает афинность (сродство) для его соединения с остальными субъединицами. Важным параметром для описания этой кривой является

42
парциальное давление О2 при половинном насыщении гемоглобина. В
человеческой крови Р0,5=27 мм рт.ст.
S-образная форма кривой связывания гемоглобином О2 имеет большое значение для транспортной функции крови. В области значений выше 65 мм
рт.ст. кривая плоская, изменение Ро2 только немного меняет насыщение кислородом. Это область нормальных значений альвеолярного Ро2, которое может немного снижаться без заметного уменьшения насыщения гемоглобина кислородом в крови капилляров легких (например при подъеме в горы).
Отвесный спад в нижней части кривой связывания (менее 65 мм рт.ст.)
гарантирует, что Ро2 в капиллярной крови периферических тканей, несмотря на отдачу кислорода, остается достаточно высоким, чтобы обеспечить ткани кислородом путем диффузии.
Ряд факторов влияет на афинность (сродство) гемоглобина к О2. При этом в первую очередь изменяется положение кривой и гораздо меньше – ее форма.
Различные метаболические факторы вызывают смещения кривой диссоциации влево или вправо (рис. 16).
ВЛЕВО ВПРАВО
Рис. 16. Сдвиги кривой диссоциации оксигемоглобина
43
Сродство гемоглобина к кислороду регулируется важнейшими факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией
2,3-дифосфоглицерата.
1) Величина рН и содержание СО2 в любой части организма закономерно изменяют сродство гемоглобина к О2: уменьшение рН крови вызывает сдвиг кривой диссоциации соответственно вправо (уменьшается сродство гемоглобина к О2), а увеличение рН крови — сдвиг кривой диссоциации влево
(повышается сродство гемоглобина к О2). Влияние рН на кривую диссоциации оксигемоглобина называется «эффектом Бора». Эффект Бора может быть связан также с изменением Рсо2: его повышение сдвигает кривую связывания О2 вправо, понижение - влево. Это влияние основывается, в первую очередь, на том, что повышение Рсо2 ведет к снижению рH и наоборот. Влияние самих молекул СО2 на афинность гемоглобина к О2, напротив, незначительно.
2)Влияние температуры: рост температуры уменьшает сродство гемоглобина к О2. В работающих мышцах увеличение температуры способствует освобождению О2. Уменьшение температуры тканей вызывает сдвиг влево кривой диссоциации оксигемоглобина. Значение влияния температуры для гомойотермных организмов небольшое, хотя температура крови на периферии тела может сильно отличаться от температуры всего организма.
3)Эффект 2,3-дифосфоглицерата: в его отсутствии афинность гемоглобина к кислороду очень высока. Главная функция 2,3-
дифосфоглицерата состоит в смещении кривой связывания О2 вправо, в
область, которая соответствует физиологическим условиям.
Контрольные вопросы
1.Какие транспортные формы кислорода существуют ?
2.Что такое кислородная емкость крови, как она определяется ?
3.Что такое коэффициент утилизации кислорода, как он определяется ?
44
4.Почему кривая диссоциации оксигемоглобина (сатурационная кривая) имеет
S- образную форму ?
5.Какое биологическое значение крутой и отлогой частей кривой диссоциации оксигемоглобина (сатурационной кривой) ?
6.Какие факторы влияют на афинность (сродство) гемоглобина к О2 ?
3.2.Транспорт углекислого газа кровью
Ввенозной крови содержится около 580 мл/л СО2. Транспорт обеспечивается в таких формах, как: 1) растворенный СО2 в плазме крови (5—
10%); 2) в виде гидрокарбонатов (80—90%); 3) карбаминовые соединения эритроцитов (5—15%).
Небольшая часть СО2 транспортируется в легкие в растворенном виде (0,3
мл/100 мл крови). Растворенный в крови СО2 реагирует с водой:
CO2 + Н2О = H2CO3
В плазме крови эта реакция протекает медленно и не имеет особого значения. Но в эритроцитах имеется цинк-содержащий фермент -
карбоангидраза - который смещает равновесие реакции вправо (в сторону образования угольной кислоты). Образование H2CO3 происходит в 1000 раз быстрее, чем в плазме, кроме того, около 99,9% H2CO3 диссоциирует с образованием HCO3-- и иона водорода (H+):
CO2 + Н2О =H2CO3 = HCO-3 + H+
Образующиеся протоны (H+) нейтрализуются гемоглобиновым буфером
(H+ + Hb = HHb). Образующийся HCO3- выходит из эритроцитов в плазму, для поддержания электронейтральности в эритроциты поступают ионы Cl -.
В эритроците CO2 может также связываться гемоглобином с образованием HbCO2. Как и в первом случае, образующийся при этом H+
связывается гемоглобиновым буфером.
Как сатурация гемоглобина кислородом коррелирует с PО2, так и общее
45
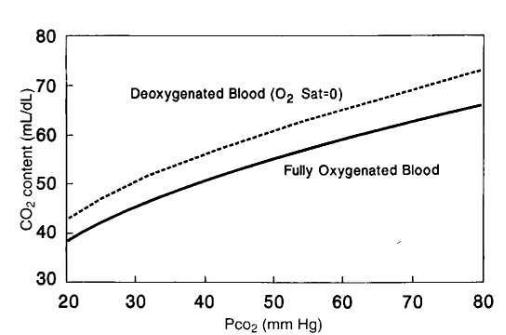
содержание CO2 в крови коррелирует с PCO2 и описывается кривой диссоциации CO2 (рис. 17).
Рис. 17. Кривая диссоциации двуокиси углерода
Даже при выраженных нарушениях Va/Q (то есть при выраженной легочной патологии) Paco2, как правило, остается в пределах нормальных значений. Это является следствием того, что кривая диссоциации CO2 (рис. 17)
нарастает монотонно. Артериовенозная разница по Pco2 в покое обычно составляет 5 мм рт. ст.и редко превышает 10 мм рт. ст. При данном значении
Pco2 деоксигенированная кровь содержит большее количество CO2, чем оксигенированная (эффект Холдена). В отличие от кривой насыщения Hb
кислородом кривая содержания CO2 не имеет плато и в клинически значимом диапазоне представляет собой прямую линию.
В венозной крови, притекающей к капиллярам легких, напряжение СО2
составляет в среднем 46 мм рт.ст., а в альвеолярном воздухе парциальное давление СО2 равно в среднем 40 мм рт.ст., что обеспечивает диффузию СО2 из плазмы крови в альвеолы легких по концентрационному градиенту.
Эндотелий капилляров проницаем только для молекулярного СО2 как полярной молекулы. Из крови в альвеолы диффундирует физически
46
растворенный в плазме крови молекулярный СО2. Кроме того, в альвеолы легких диффундирует СО2, который высвобождается из карбаминовых соединений эритроцитов благодаря реакции окисления гемоглобина в капиллярах легкого, а также из гидрокарбонатов плазмы крови в результате их быстрой диссоциации с помощью фермента карбоангидразы, содержащейся в эритроцитах. Молекулярный СО2 проходит аэрогематический барьер, а затем поступает в альвеолы. В норме через 1 с происходит выравнивание концентраций СО2 на альвеолярно-капиллярной мембране, поэтому за половину времени капиллярного кровотока происходит полный обмен СО2
через аэрогематический барьер. Реально равновесие наступает несколько медленнее. Это связано с тем, что перенос СО2, так же как и О2, ограничивается скоростью перфузии капилляров легких.
Контрольные вопросы
1.Какие транспортные формы углекислого газа существуют ?
2.Какая форма транспорта углекислого газа основная ?
3.Почему кривая диссоциации двуокиси углерода в виде прямой линии ?
3.3. Тестовые задания и ситуационная задача
Выберите один правильный ответ.
20.УГЛЕКИСЛЫЙ ГАЗ В КРОВИ ТРАНСПОРТИРУЕТСЯ В ВИДЕ
1)растворенного газа, связанного с гемоглобином, в составе бикарбонатов
2)только в растворенном виде
3)только в составе бикарбонатов
21.ДИСССОЦИАЦИЯ ОКСИГЕМОГЛОБИНА УВЕЛИЧИВАЕТСЯ ПРИ УСЛОВИИ
1)увеличения pH крови, уменьшения содержания CO2 в крови
2)уменьшения температуры тела, увеличения содержания карбоксигемоглобина
3)увеличения pH крови, повышения температуры тела, увеличения содержания CO2 в крови
4)увеличения pH крови и повышения температуры тела
47
22.КИСЛОРОДНАЯ ЕМКОСТЬ КРОВИ – ЭТО
1)максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом
2)диффузия кислорода и альвеолярного воздуха в кровь
3)часть кислорода, поглощаемого тканями из венозной крови
4)часть кислорода, поглощаемого тканями из артериальной крови
23.КОЭФФИЦИЕНТОМ УТИЛИЗАЦИИ КИСЛОРОДА НАЗЫВАЮТ
1)максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом
2)большую часть кислорода, содержащуюся в артериальной крови
3)часть кислорода, поглощённую тканями из венозной крови
4)часть кислорода, поглощённую тканями из артериальной крови
Ситуационная задача 3
У больного резко нарушен транспорт гемоглобина. Какое терапевтическое воздействие может помочь в обеспечении тканей кислородом ?
4. Газообмен в тканях
Обмен СО2 между клетками тканей с кровью тканевых капилляров определяется следующими механизмами. В процессах окисления в тканях образуется СО2. Обычно в большинстве тканей рСО2 близок в 50-60 мм рт. ст. В
крови, поступающей в артериальный конец капилляров раСО2= 40 мм рт.ст.
Наличие градиента заставляет СО2 диффундировать из тканевой жидкости к капиллярам. Интенсивность окисления в различных тканях не одинакова.
Поэтому в смешанной венозной крови, поступающей в правое предсердие в покое, рvcо2 равно 46 мм рт.ст. Кровь, проходящая через легкие отдает не весь СО2. Большая часть его сохраняется в артериальной крови.
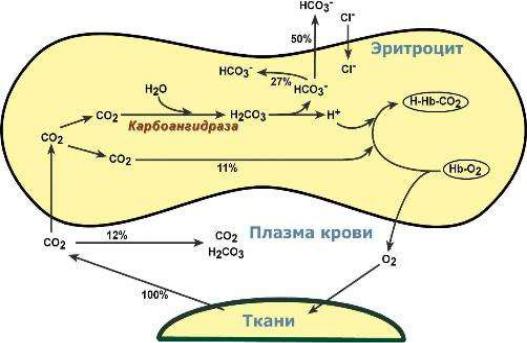
В ходе газообмена СО2 между тканями и кровью содержание НСО3- в
эритроците повышается и они начинают диффундировать в кровь (рис. 18). Для поддержания электронейтральности в эритроциты начнут поступать из плазмы дополнительно ионы С1-. Наибольшее количество бикарбонатов плазмы крови
48
образуется при участии карбоангидразы эритроцитов. В условиях in vitro
образование молекулярного СО2 из гидрокарбонатов происходит чрезвычайно медленно и диффузия этого газа занимает около 5 мин, тогда как в капиллярах легкого равновесие наступает через 1 с. Это определяется функцией фермента карбоангидразы угольной кислоты. В функции карбоангидразы выделяют следующие типы реакций:
СО2+Н2О =H2СО-3 = H++НСО3-
Рис. 18. Газообмен в тканях
Карбаминовый комплекс СО2 с гемоглобином образуется в результате реакции СО2 с радикалом NH2 глобина. Эта реакция протекает без участия какого-либо фермента, т. е. она не нуждается в катализе. Реакция СО2 с Нb
приводит, во-первых, к высвобождению Н+; во-вторых, в ходе образования карбаминовых комплексов снижается сродство Нb к О2.
Контрольные вопросы
1.Чему равно напряжение углекислого газа в тканях, артериальной и венозной крови ?
49
2.Какова последовательность газообмена в тканях ?
3.Какой тип реакции обеспечивает карбоангидраза ?
4.1. Тестовые задания
Выберите один правильный ответ.
24.ФЕРМЕНТ КАРБОАНГИДРАЗА НАХОДИТСЯ
1)в слизи
2)в сурфактанте
3)в эритроцитах
4)в плазме крови
25.НАПРЯЖЕНИЕ УГЛЕКИСЛОГО ГАЗА В ВЕНОЗНОЙ КРОВИ СОСТАВЛЯЕТ
1)100 мм рт.ст.
2)46 мм рт.ст.
3)40 мм рт.ст.
4)15 мм рт.ст.
5. Тканевое дыхание
Более 90% всей энергии в организме образуется в результате горения водорода. Сам данный процесс суммарно можно представить в ходе простой химической реакции:
2Н2 + О2  2Н2О Термин «горение» применительно к организму применим весьма условно,
2Н2О Термин «горение» применительно к организму применим весьма условно,
хотя суть процесса реакция отображает правильно. Реакция взаимодействия водорода с кислородом с химической точки зрения является окислительно-
восстановительной, поэтому применительно к организму используется другой термин - биологическое окисление. Водород используется как главное топливо для образования энергии. В митохондриях поток электронов от водорода устремляется к их главному акцептору кислороду. При этом образуются молекулы воды, которая в энергетической шкале биологических веществ
50
занимает низшую ступеньку и является конечным продуктом тканевого дыхания. Предыдущие этапы дыхания обеспечивают лишь доставку кислорода к клеткам, где в митохондриях функционирует цепь переноса электронов
(дыхательная цепь). При этом происходит перенос по цепи переносчиков вначале водорода, а затем электронов на кислород, с постепенным освобождением энергии, значительная часть которой аккумулируется в макроэргических связях АТФ. У теплокровных животных, в том числе и у человека, работа дыхательной цепи одновременно обеспечивает выработку тепла, которое идет на поддержание температуры тела.
Процесс тканевого дыхания носит достаточно сложный и многоступенчатый характер, начало изучения, которого было положено в XVIII веке. Следует отметить, что работы ученых, о вкладе которых в изучения процессов биологического окисления и тканевого дыхания будет говориться ниже, не потеряли значения до настоящего времени.
5.1. История изучения процессов тканевого дыхания
Первые представления о тканевом дыхании связаны с Лавуазье, который один из первых указал на то, что жизнь поддерживается кислородом. Он назвал дыхание процессом биологического «горения», подобным горению угля, только очень медленным. Таким образом, Лавуазье были обнаружены сходство и отличия между процессами горения и биологического окисления, которые заключались в следующем:
Сходства:
1.Как горение, так и биологическое окисление идут с потреблением кислорода;
2.Конечными продуктами горения и биологического окисления являются вода
и углекислый газ.
Отличия биологического окисления от горения заключаются в следующем: