

2 курс / Нормальная физиология / ФЗЛ ЧЕЛОВЕКА
.pdfВ ответ на начальную деполяризацию мембраны, вызванную раздражителем, открывается лишь небольшое число натриевых каналов. Их открывание, однако, приводит к возникновению входящего внутрь клетки потока ионов Na+ (входящий натриевый ток), который увеличивает начальную деполяризацию. Это ведет к открыванию новых натриевых каналов, т. е. к дальнейшему повышению gNa соответственно входящего натриевого тока, а следовательно, к дальнейшей деполяризации мембраны, что, в свою очередь, обусловливает еще большее повышение gNa и т. д. Такой круговой лавинообразный процесс получил название регенеративной (т. е. самообновляющейся) деполяризации.
Схематически он может быть изображен следующим образом:
Теоретически регенеративная деполяризация должна была бы завершаться повышением внутреннего потенциала клетки до величины равновесного нернстовского потенциала для ионов Na+:
где Na0+— наружная, а Nai+ — внутренняя концентрация ионов Na+,
При наблюдаемом соотношении
Эта величина является предельной для потенциала действия. В действительности, однако, пиковый потенциал никогда не достигает величины ENa, во-первых, потому, что мембрана в момент пика потенциала действия проницаема не только для, ионов Na+, но и для ионов К+ (в значительно меньшей степени). Во-вторых, подъему потенциала действия до величины ENa противодействуют восстановительные процессы, ведущие к восстановлению исходной поляризации (реполяризация мембраны).
Такими процессами являются снижение значения gNa и повышение уровня gK- Снижение gNa обусловлено тем, что активация натриевых каналов во время деполя-
ризации сменяется их инактивацией; это приводит к быстрому уменьшению числа открытых натриевых каналов. Одновременно под влиянием деполяризации начинается медленная активация калиевых каналов, обусловливающая рост значения gk. Следствием увеличения gK является усиление выходящего из клетки потока ионов К+ (выходящий калиевый ток).
В условиях понижения gNa, связанного с инактивацией натриевых каналов, выходящий ток ионов К+ приводит к реполяризации мембраны или даже к ее временной («следовой») гиперполяризации, как это имеет место, например, в гигантском аксоне кальмара
(см. рис. 4).
Реполяризация мембраны в свою очередь ведет к закрыванию калиевых каналов и, следовательно, ослаблению выходящего калиевого тока. Вместе с тем под влиянием реполяризации происходит медленное устранение натриевой инактивации:
открываются инактивационныё ворота и натриевые каналы возвращаются в состояние покоя.
На рис. 9 схематически показано состояние натриевых и калиевых каналов в различные фазы развития потенциала действия.
Все агенты, блокирующие натриевые каналы (тетродотоксин, местные анестетики и многие другие препараты), снижают крутизну нарастания и амплитуду потенциала действия и тем в большей степени, чем выше концентрация этих веществ.
3J

АКТИВАЦИЯ НАТРИЙ-КАЛИЕВОГО НАСОСА ПРИ ВОЗБУЖДЕНИИ
Возникновение серии импульсов в нервном или мышечном волокне сопровождается обогащением протоплазмы Na+ и потерей К+. Для гигантского аксона кальмара диаметром 0,5 мм подсчитано, что во время одиночного нервного импульса через каждый квадратный микрон мембраны в протоплазму поступает около 20 000 Na+ и столько же К+ покидает волокно. В итоге при каждом импульсе аксон теряет около одной миллионной общего содержания калия. Хотя эти потери очень незначительны, при ритмическом следовании импульсов, суммируясь, они должны были бы привести к более или менее заметным изменениям концентрационных градиентов.
Особенно быстро такие концентрационные сдвиги должны были бы развиваться в тонких нервных и мышечных волокнах и мелких нервных клетках, обладающих малым по отношению к поверхности объемом цитоплазмы. Этому, однако, противодействует натриевый насос, активность которого возрастает при повышении внутриклеточной концентрации ионов Na+.
Усиление работы насоса сопровождается значительным повышением интенсивности обменных процессов, поставляющих энергию для активного переноса ионов Na+ и К+ через мембрану. Это проявляется усилением процессов распада и синтеза АТФ и креатинфосфата, увеличением потреблении клеткой кислорода, повышением теплопродукции и т. п.
Благодаря работе насоса нарушенное при возбуждении неравенство концентраций Na+ и К+ по обе стороны мембраны полностью восстанавливается. Следует, однако, подчеркнуть, что скорость выведения Na+ из цитоплазмы с помощью насоса относительно мала: она примерно в 200 раз ниже скорости движения этих ионов через мембрану по концентрационному градиенту.
Таким образом, в живой клетке существует две системы движения ионов через мембрану (рис. 10). Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии, поэтому его называют пассивным ионным транспортом. Он ответствен за возникновение потенциала покоя и потенциала действия и ведет в конечном итоге к выравниванию концентрации ионов по обе стороны клеточной мембраны. Второй тип движения ионов через мембрану, осуществляющийся против концентрационного градиента, состоит в «выкачивании» ионов натрия из цитоплазмы и «нагнетании» ионов калия внутрь клетки. Этот тип ионного транспорта возможен лишь при условии затраты энергии обмена веществ. Его называют активным ионным транспортом. Он ответствен за поддержание постоянства разности концентраций ионов между цитоплазмой и омывающей клетку жидкостью. Активный транспорт — результат работы натриевого насоса, благодаря которому восстанавливается исходная разность ионных концентраций, нарушающаяся при каждой вспышке возбуждения.
32
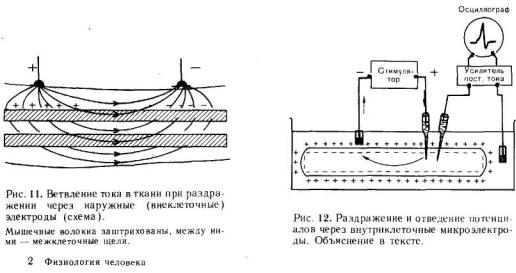
МЕХАНИЗМ РАЗДРАЖЕНИЯ КЛЕТКИ (ВОЛ ОКНА) ЭЛЕКТРИЧЕСКИМ ТОКОМ
В естественных условиях генерацию потенциала действия вызывают так называемые местные токи, возникающие между возбужденным (деполяризованным) и покоящимся участками клеточной мембраны. Поэтому электрический ток рассматривается как адекватный раздражитель для возбудимых мембран и успешно используется в экспериментах при изучении закономерностей возникновения потенциалов действия.
Минимальную силу тока, необходимую и достаточную для инициации потенциала действия, называют пороговой, соответственно раздражители большей и меньшей силы обозначают подпороговыми и сверхпороговыми. Пороговая сила тока (пороговый ток) в определенных пределах находится в обратной зависимости от длительности его действия. Существует также некоторая минимальная крутизна нарастания силы тока, ниже которой последний утрачивает способность вызывать потенциал действия.
Существуют два способа подведения тока к тканям для измерения порога раздражения и, следовательно, для определения их возбудимости. При первом способе — внеклеточном — оба электрода располагают на поверхности раздражаемой ткани. Условно принимают, что приложенный ток входит в ткань в области анода и выходит в области катода (рис. 11). Недостаток этого метода измерения порога заключается в значительном ветвлении тока: только часть его проходит через мембраны клеток, часть же ответвляется в межклеточные щели. Вследствие этого при раздражении приходится применять ток значительно большей силы, чем необходимо для возникновения возбуждения.
При втором способе подведения тока к клеткам — внутриклеточном — микроэлектрод вводят в клетку, а обычный электрод прикладывают к поверхности ткани (рис. 12). В этом случае весь ток проходит через мембрану клетки, что позволяет точно определить наименьшую силу тока, необходимую для возникновения потенциала действия. При таком способе раздражения отведение потенциалов производят с помощью второго внутриклеточного микроэлектрода.
Пороговая сила тока, необходимая для возникновения возбуждения различных клеток при внутриклеточном раздражающем электроде, равна 10-7— 10- 9 А.
В лабораторных условиях и при проведении некоторых клинических исследований для раздражения нервов и мышц применяют электрические стимулы различной формы: прямоугольной, синусоидальной, линейно и экспоненциально нарастающей, индукционные удары, конденсаторные разряды и т. п.
Механизм раздражающего действия тока при всех видах стимулов в принципе одинаков, однако в наиболее отчетливой форме он выявляется при использовании постоянного тока.
33
ДЕЙСТВИЕ ПОСТОЯННОГО ТОКА НА ВОЗБУДИМЫЕ ТКАНИ
Полярный закон раздражения
При раздражении нерва или мышцы постоянным током возбуждение возникает в момент замыкания постоянного тока только под катодом, а в момент размыкания — только под анодом. Эти факты объединяют под названием полярного закона раздражения, открытого Пфлюгером в 1859 г. Полярный закон доказывается следующими опытами. Умерщвляют участок нерва под одним из электродов, а второй электрод устанавливают на неповрежденном участке. Если с неповрежденным участком соприкасается катод, возбуждение возникает в момент замыкания тока; если же катод устанавливают на поврежденном участке, а анод — на неповрежденном, возбуждение возникает только при размыкании тока. Порог раздражения при размыкании, когда возбуждение возникает под анодом, значительно выше, чем при замыкании, когда возбуждение возникает под катодом.
Изучение механизма полярного действия электрического тока стало возможным только после того, как был разработан описанный метод одновременного введения в
клетки двух микроэлектродов: одного — для раздражения, другого — для отведения потенциалов. Было установлено, что потенциал действия возникает только в том случае, если катод находится снаружи, а анод — внутри клетки. При обратном расположении полюсов, т. е. наружном аноде и внутреннем катоде, возбуждения при замыкании тока не возникает, как бы силен он ни был.
Прохождение через нервное или мышечное волокно электрического тока прежде всего вызывает изменения мембранного потенциала.
В области приложения к поверхности ткани анода положительный потенциал на наружной стороне мембраны возрастает, т. е. происходит гиперполяризация, а в том случае, когда к поверхности приложен катод, положительный потенциал на наружной стороне мембраны снижается — возникает деполяризация.
На рис. 13, а показано, что как при замыкании, так и при размыкании тока изменения мембранного потенциала нервного волокна не возникают и не исчезают мгновенно, а плавно развиваются во времени.
Объясняется это тем, что поверхностная мембрана живой клетки обладает свойствами конденсатора. Обкладками этого «тканевого конденсатора» служат наружная и внутренняя поверхности мембраны, а диэлектриком — слой липидов, обладающий значительным сопротивлением. Ввиду наличия в мембране каналов, через которые могут проходить ионы, сопротивление этого слоя не равно бесконечности, как в идеальном конденсаторе. Поэтому поверхностную мембрану клетки обычно уподобляют конденсатору с параллельно включенным сопротивлением, по которому может происходить утечка зарядов (рис. 13,а).
Временной ход изменений мембранного потенциала при включении и выключении тока (рис. 13, б) зависит от емкости С и сопротивления мембраны R. Чем меньше произведение RC — постоянная времени мембраны, тем быстрее при данной силе тока нарастает потенциал и, наоборот, большей величине RC соответствует меньшая скорость увеличения потенциала.
Изменения мембранного потенциала возникают не только непосредственно в точках приложения к нервному волокну катода и анода постоянного тока, но и на некотором расстоянии от полюсов с той, однако, разницей, что их величина постепенно убывает по мере удаления от катода и анода. Объясняется это так называемыми кабельными свойствами нервного и мышечного волокон. Однородное нервное волокно в электрическом отношении представляет собой кабель, т. е. сердечник с низким удельным сопротивлением (аксоплазма), покрытый изоляцией (мембраной) и помещенный в хорошо проводящую среду. Эквивалентная схема кабеля приведена на рис. 13, б. При пропускании через некоторую точку волокна длительное время постоянного тока наблюдается стационарное состояние, при котором плотность тока и, следовательно, изменение мембранного потенциала максимальны в месте приложения тока (т. е. непосредственно под катодом и анодом); с удалением от полюсов плотность тока и изменения потенциала на мембране экспоненциально уменьшаются по длине волокна. Поскольку рассматриваемые изменения мембранного потенциала в отличие от локального ответа потенциала действия или следовых потенциалов не связаны с изменениями ионной проницаемости мембраны (т. е. активным ответом волокна), их принято называть пассивными,
34
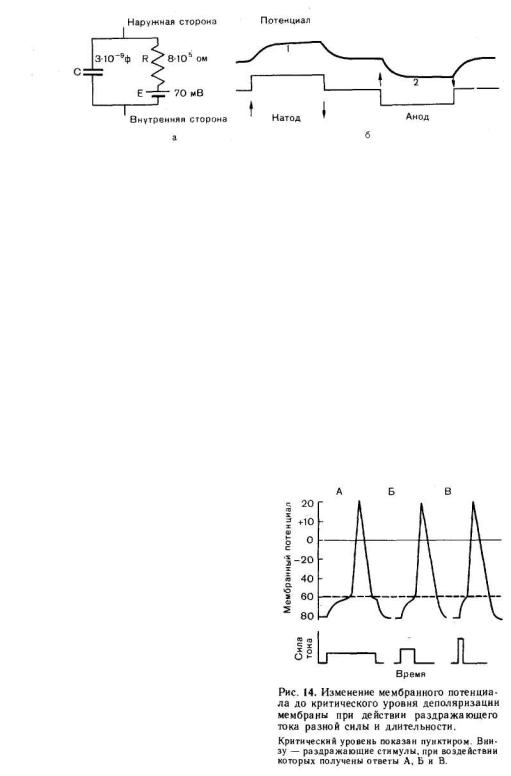
Рис. 13. Простейшая электрическая схема, воспроизводящая электрические свойства мембраны (а) и изменения мембранного потенциала под катодом и анодом постоянного тока подпороговой силы (б).
а: С — емкость мембраны, R — сопротивление, Е — электродвижущая сила мембраны в покое (п отенциал покоя). Приведены средние значения R, С и Е для мотонейрона, б — деполяризация мембраны (l) под катодом и гиперполяризация (2) под анодом при прохождении через нервное волокно слабого подпорогового тока.
или «электротоническими», изменениями мембранного потенциала. В чистом виде последние могут быть зарегистрированы в условиях полной блокады ионных каналов химическими агентами. Различают кат- и анэлектротонические изменения потенциала, развивающиеся в области приложения соответственно катода и анода постоянного тока.
Критический уровень деполяризации
Регистрация изменений мембранного потенциала при внутриклеточном раздражении нервного или мышечного волокна показала, что потенциал действия возникает в тот момент, когда деполяризация мембраны достигает критического уровня. Этот критический уровень деполяризации не зависит от характера примененного стимула, расстояния между электродами и т. п., а определяется исключительно свойствами самой мембраны.
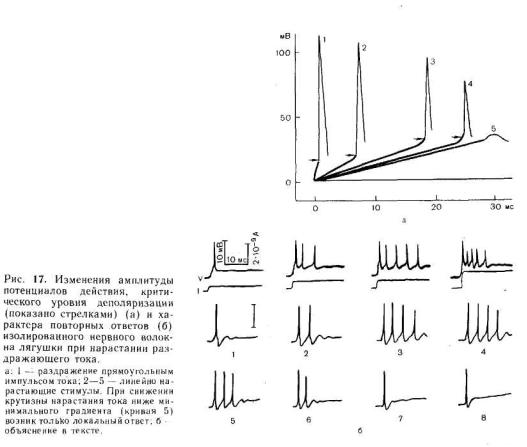
На рис. 14 схематически показаны изменения мембранного потенциала нервного волокна под влиянием длительного и_коротких стимулов различной силы. Во всех случаях потенциал действия возникает тогда, когда мембранный потенциал достигает критической величины. Скорость, с которой происходит деполяризация мембраны, при прочих равных условиях зависит от силы раздражающего тока. При токе слабой силы деполяризация развивается медленно, поэтому для возникновения потенциала действия стимул должен быть большей длительности. В случае усиления раздражающего тока скорость развития деполяризации возрастает и соответственно уменьшается минимальное время, необходимое для возникновения возбуждения. Чем быстрее развивается деполяризация мембраны, тем меньше минимальное время, необходимое для генерации потенциала действия, и наоборот.
Локальный ответ
В механизме критической деполяризации мембраны наряду с пассивными существенную роль играют активные подпороговые изменения мембранного потенциала, проявляющиеся в форме так называемого локального ответа.

Первые признаки локального ответа появляются при действии стимулов, составляющих 50—75 % от пороговой величины. По мере дальнейшего усиления раздражающего тока локальный ответ увеличивается, и в момент, когда деполяризация мембраны, обусловленная суммой катэлектротонического потенциала и локального ответа, достигает критического уровня, возникает потенциал действия (рис. 15).
Локальный ответ, так же как и потенциал действия, обусловлен повышением натриевой проницаемости мембраны. Однако при подпороговом стимуле это начальное повышение натриевой проницаемости недостаточно велико, чтобы вызвать быструю регенеративную деполяризацию мембраны. Развитие деполяризации тормозится процессами инактивации натриевых и активации калиевых каналов. Поэтому рост локального ответа приостанавливается, а затем происходит реполяризация мембраны. Амплитуда локального ответа увеличивается по мере приближения силы стимула к порогу, и при достижении последнего локальный ответ перерастает в потенциал действия, поскольку скорость увеличения натриевой проницаемости мембраны начинает превышать скорость роста калиевой проницаемости.
Зависимость пороговой силы раздражителя от его длительности
Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Особенно четко эта зависимость проявляется при использовании в качестве раздражителя прямоугольных импульсов постоянного тока.
Представленная на рис. 16 кривая называется кривой силы — длительности, или силы — времени. Она была изучена при исследовании различных нервов и мышц Гоорве-
гом (1892), Вейсом (1901) и Лапиком (1909).
По этой кривой прежде всего можно судить о том, что ток ниже некоторой минимальной силы или напряжения не вызывает возбуждения, как бы длительно он ни действовал. Минимальная сила постоянного тока, способная вызвать возбуждение (порог раздражения), названа Лапиком реобазой (ордината ОА). Наименьшее время (отрезок ОС), в течение которого должен действовать раздражающий стимул, величиной в одну реобазу называют полезным временем. Слово «полезное» здесь применено с целью подчеркнуть, что дальнейшее увеличение длительности действия тока не имеет значения (бесполезно) для возникновения потенциала действия.
Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. Как видно на рис. 16, при очень коротких стимулах кривая силы — времени становится параллельной оси ординат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражителя. Поэтому, кроме полезного времени, в качестве времени константы раздраже-
36
ния Лапик ввел понятие «хронаксия». Хронаксия - это время, в течение которого должен действовать ток удвоенной реобазы, чтобы вызвать возбуждение.
В настоящее время показано, что величина хронаксии зависит от RC мембраны и временной константы процесса активации натриевых (или кальциевых) каналов. Точное измерение величин реобазы или хронаксии возможно только в опытах на одиночных возбудимых клетках. При раздражении целой мышцы или нерва (особенно через кожу, как это делается при исследованиях возбудимости, проводимых на человеке) ветвление тока и поляризация окружающих тканей вносят очень большие искажения в измеряемые величины. Все же в некоторых случаях использование хронаксиметрии оказалось полезным в неврологической практике: с ее помощью удается установить наличие органического поражения (перерождения) двигательного нерва. Дело в том, что электрический ток, приложенный к мышце, проходит и через находящиеся в ней нервные волокна и их окончания. Величины реобазы и хронаксии нервных волокон значительно меньше соответствующих величин мышечных волокон, поэтому при пороговых силах тока возбуждение прежде всего возникает в нервных волокнах и от них передается на мышцу. Из этого следует, что при измерении хронаксии мышцы фактически получают значение хронаксии иннервирующих ее нервных волокон. Если нерв поврежден или произошла гибель соответствующих мотонейронов в спинном мозге (как это, например, имеет место при полиомиелите), то нервные волокна перерождаются, тогда раздражающий стимул выявляет хронаксию собственно мышечных волокон, которая имеет большую продолжительность.
Явление аккомодации
Пороговая сила тока увеличивается при уменьшении крутизны его нарастания, а при некоторой минимальной крутизне ответы на раздражение исчезают. Это явление принято обозначать термином «аккомодация». На рис. 17, а показаны изменения критического уровня деполяризации и амплитуды потенциала действия при раздражении одиночного нервного волокна лягушки линейно нарастающими токами различной крутизны. Уменьшение последней приводит к повышению критического уровня деполяризации (примерно на 20 % от исходной величины) и снижению амплитуды потенциалов

действия. При снижении крутизны до некоторого минимального уровня («минимальный градиент», или «критический наклон») потенциал действия не возникает. Величина этого «минимального градиента», выраженного в единицах реобаза в секунду, принята
вкачестве меры скорости аккомодации.
Воснове аккомодации лежат инактивация натриевой и повышение калиевой проводимостей, развивающиеся во время медленно нарастающей деполяризации мем-- браны. Аккомодация различных нервных волокон варьирует в широких пределах, но у двигательных нервных волокон скорость аккомодации, как правило, значительно вы-
ше, чем у чувствительных волокон.
Повторные ответы
Многие нервные волокна обладают способностью отвечать несколькими потенциалами действия на включение постоянного деполяризующего тока. Эта способность к повторным ответам особенно хорошо выражена у сенсорных волокон, для которых длительная деполяризация мембраны рецептора является естественным раздражителем. Примеры повторных ответов на постоянный ток представлены на рис. 17, б. Как правило, пороговая сила тока, необходимая для возникновения повторных ответов, выше, чем для инициации одиночного потенциала действия. На рисунке видно, что увеличение силы деполяризующего тока до определенной величины обусловливает возрастание частоты импульсов и увеличение их числа (1—4). Однако при дальнейшем повышении силы тока частота импульсов уменьшается и в конечном итоге возникает только одиночный потенциал действия (5—8).
При постоянной силе тока длина межимпульсных интервалов в повторном ответе постепенно увеличивается. Это явление получило название адаптации. В его основе лежит медленное повышение калиевой проводимости мембраны, связанное с активацией особых медленных калиевых каналов. Эти каналы найдены как в нервных волокнах, так и в нервных клетках, у которых способность к повторным ответам и явление адаптации, как правило, хорошо выражены.
Изменение критического уровня деполяризации
Было показано, что условием возникновения потенциала действия является критическая деполяризация мембраны. Если исходный потенциал мембраны перед нанесением раздражающего стимула обозначить £о, а критическую величину мембранного потенциала Ek. то указанное условие порогового раздражения можно записать так:
— пороговый потенциал, т. е. величина, на которую необходимо повысить внутренний потенциал мембраны для возникновения импульса.
Эта простая зависимость помогла понять причину известных со времени классических исследований Пфлюгера (1859) изменений возбудимости нервного волокна в области катода и анода постоянного тока. В области катода мембрана деполяризуется, т. е. значение E0 приближается к величине Eк, соответственно уменьшается и, следовательно, возбудимость возрастает. В области анода, напротив, E0 уменьшается
(внутренний потенциал мембраны становится более отрицательным); теперь для критической деполяризации мембраны необходимо ее потенциал сместить на большую величину —возбудимость снижается (рис. 18).
При длительной деполяризации мембраны развиваются процессы, повышающие критический уровень деполяризации. Такими процессами являются инактивация натриевых каналов и активация калиевых. Рост Eк при данном значении E0 ведет к увеличению порогового потенциала, т .е . снижению возбудимости (рис. 19). Одновременно с увеличением порога происходит снижение потенциала действия — крутизна его нарастания и амплитуда падают. Такое снижение возбудимости нервного волокна при длительной и сильной деполяризации мембраны было впервые описано Вериго (1889)
и получило название катодической депрессии.
38
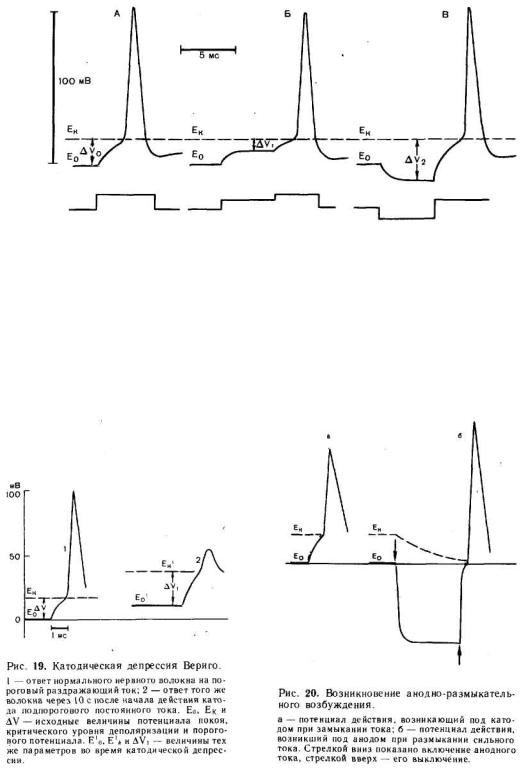
Рис. 18. Соотношения между исходным уровнем потенциала покоя ( Е о ) , критическим уровнем деполяри зации (Е к ) и п ороговым п отен циалом (∆V), в н орме (А) и п ри элек тротон е (Б, В ) .
Б — катэлсктротоническое снижение порога ∆Vi наступает в результате приближения потенциала покоя Ео к критическому уровню деполяризации Ек; В — анэлектротоническое повышение порога ∆V2 является следствием удаления исходного уровня потенциала покоя Ео от Ек. Нижняя линия — раздражающий и поляризующий токи: катодный ток — вверх, анодный ток — вниз.
Описанные явления демонстрируют наблюдающиеся в естественных условиях изменения возбудимости нервных клеток и пресинаптическйх нервных терминалей при действии на них медиаторов, вызывающих кратковременную или длительную деполяризацию мембраны. В частности, предполагают, что явление катодической депрессии лежит в основе так называемого пресинаптического торможения, наблюдающегося в ЦНС.
39
В заключение необходимо рассмотреть изменения возбудимости, наблюдающиеся при длительной гиперполяризации мембраны. Такая гиперполяризация (в нервном волокне она появляется в области приложения анода) приводит к снижению калиевой проницаемости и ослаблению исходной натриевой инактивации. Эти изменения ведут к снижению ∆V возросшего в начальный момент гиперполяризации мембраны, и увеличению амплитуды и крутизны нарастания потенциала действия. Если гиперполяризующий ток прикладывается к мембране, которая предварительно была подвергнута воздействию избытка ионов К+ или действию анестетиков, усиливающих исходную натриевую инактивацию (т. е. увеличивающих долю каналов, у которых инактивационные «ворота» закрыты), то во время длительной гиперполяризации ∆V может снизиться настолько, что при выключении тока, т. е. при возвращении мембранного потенциала к его исходной величине, возникает потенциал действия (рис. 20). Такое явление получило название анодно-размыкательного возбуждения. Предполагают, что в некоторых нервных клетках на подобном механизме основано возникновение потенциала действия после окончания тормозного гиперполяризационного потенциала в естественных условиях. По-видимому, у таких клеток инактивация натриевых каналов и активация калиевых сильно выражены при потенциале покоя в нормальной солевой среде.
Изложенные данные о влиянии деполяризации и гиперполяризации мембраны на ее возбудимость представлены в табл. 2.
Физиологические |
Деполяризация мембраны |
Гиперполяризация мембраны |
|||
параметры |
(к атэлектротон) |
|
(а нэлектротон) |
||
|
|
|
|||
Возбудимость |
Вначале повышение, затем пони- |
Сначала понижение, затем относи- |
|||
Потенциал действия |
жение |
прогрессирующее во |
тельное повышение |
||
Понижение, |
Повышение, прогрессирующее во |
||||
|
времени |
вплоть |
до |
полного |
времени |
Скорость проведения |
угнетения |
|
|
|
Вначале понижение вплоть до бло- |
Вначале повышение, затем пони- |
|||||
|
жение вплоть до полного блока |
ка (при сильном токе), затем |
|||
Натриевая |
Вначале повышение, приводящее |
постепенное восстановление |
|||
Постепенное ослабление инактива- |
|||||
мость |
к возникновению при подпорого- |
ции, если она имела место |
|||
|
вом токе |
локального |
ответа, |
|
|
|
а при пороговом — потенциала |
|
|||
|
действия, |
затем |
постепенная |
|
|
Калиевая проницаемость |
инактивация |
|
|
Понижение, если она была по- |
|
Постепенное повышение |
|
||||
|
|
|
|
|
вышена |
|
|
|
|
|
|
ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ
Используя тестирующие стимулы в различные фазы развития потенциала действия, можно проследить временной ход изменений возбудимости, сопровождающих возбуждение. На рис. 21 видно, что во время развития локального ответа возбудимость повышается (мембранный потенциал приближается к критическому уровню деполяризации); во время пика потенциала действия мембрана утрачивает возбудимость, которая постепенно восстанавливается после окончания пика. Период полной невозбудимости получил название «фаза абсолютной рефрактерности». Она обусловлена практически полной инактивацией натриевых каналов и повышением калиевой проводимости. Реполяризация мембраны ведет к реактивации натриевых каналов и
40