

2 курс / Нормальная физиология / Основы_физиологии_человека_Том_1_Агаджанян_Н_А_ред_2012
.pdfсочной, лежит островковая доля (insula). На медиальной части полушария, на границе со стволом мозга находится лимбическая доля. На нижней поверхности мозга располагается обонятельная кора. Оба полушария соединены мозолистым телом (corpus collosum).
Филогенетически кору больших полушарий делят на
древнюю (архикортекс), старую (палеокортекс) и новую ко-
ру (неокортекс). К архикортексу относят обонятельные луковицы, обонятельные тракты, расположенные на нижней поверхности лобной доли, обонятельные бугорки с обонятельными центрами. Палеокортекс включает поясную извилину, гиппокамп и миндалину. Все остальные области относят к неокортексу, который особенно хорошо развит у млекопитающих и человека.
В коре различают три основных типа нейронов. Пирамидные клетки, имеющие длинные аксоны, заходящие в другие отделы мозга, и дендриты, покрытые большим количеством шипиков – синаптических структур, благодаря которым нервная клетка контактирует с другими нервными элементами. Шипики крайне чувствительны к различным факторам: гипоксии, асфиксии, влиянию токсических веществ, под действием которых они атрофируются и при этом нарушаются функциональные связи.
Звездчатые клетки имеют короткие дендриты и аксон, их функция сводится к обеспечению связей между нейронами самой коры.
Веретенообразные клетки образуют вертикальные или горизонтальные связи нейронов разных слоев коры.
Неокортекс имеет шестислойное строение (рис. 3.16).
Первый слой – молекулярный, или плексиформный, со-
держит мало клеток, но в основном нервные волокна, образованные восходящими дендритами пирамидных нейронов и волокнами неспецифических ядер зрительного бугра, регулирующих уровень возбудимости коры больших полушарий.
169
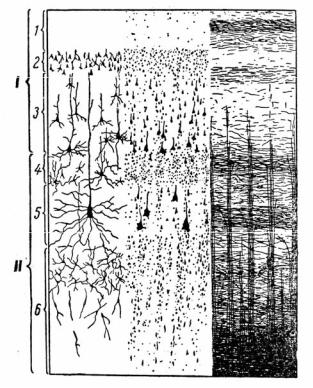
Рис. 3.16. Строение коры большого мозга:
1 – молекулярный, или плексиформный, слой; 2 – наружный гранулярный, или зернистый, слой; 3 – наружный пирамидный слой; 4 – внутренний зернистый слой; 5 – внутренний пирамидный слой; 6 – мультиформный слой
Второй слой – наружный зернистый, или наружный гранулярный, состоит из звездчатых клеток, волокна которого располагаются в основном вдоль поверхности коры и образуют кортико-кортикальные связи, ответственные за циркуляцию возбуждения в коре, т.е. кратковременную память.
Третий слой – наружный пирамидный, состоит в ос-
новном из пирамидных клеток малой и средней величины и совместно со вторым слоем обеспечивает «межкорковую» передачу информации.
170
Четвертый слой – внутренний зернистый, содержит звездчатые клетки, на которых заканчиваются специфические таламокортикальные афферентные пути от анализаторов.
Пятый слой – внутренний слой крупных пирамидных клеток, аксоны которых спускаются в продолговатый и спинной мозг. В моторной коре в этом слое находятся гигантские пирамидные клетки Беца, апикальные дендриты которых достигают поверхностных слоев, а самые длинные аксоны образуют пирамидный тракт, достигающий спинного мозга.
Шестой слой – мультиморфные клетки веретенооб-
разной и треугольной формы, образующие кортикоталамические пути.
Таким образом, каждый слой коры выполняет определенную функцию. В первом и четвертом слоях происходит восприятие и обработка поступившей в кору информации. Второй и третий слои обеспечивают кортико-кортикальные ассоциативные связи. В пятом и шестом слоях формируются эфферентные пути, идущие из коры к нижележащим структурам мозга.
Распределение нейронов различается в определенных областях коры. Это позволило Бродману выделить 52 цитоархитектонических поля (рис. 3.17). Кроме того, по функциональным признакам нейроны коры объединяются в специальные модули – «колонки», расположенные вертикально. Каждая колонка отвечает за определенный вид информации. Она связана с соседними колонками реципрокными отношениями – возбуждение одной приводит к торможению соседних. Так, например, каждая колонка связана с определенным суставом и от нее идет команда к мышцам.
Возбуждающим медиатором пирамидных клеток может быть или глутамат, или аспартат, тормозным – ГАМК.
Часть нейронов выделяет моноамины: норадреналин и дофа-
мин, другая – ацетилхолин.
171
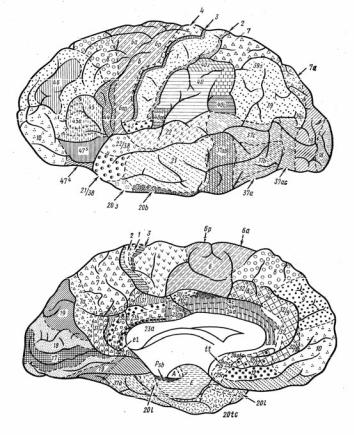
Рис. 3.17. Цитоархитектонические поля большого мозга по Бродману:
вверху – наружная поверхность полушария; внизу – внутренняя поверхность полушария
Локализация функций в коре больших полушарий.
В настоящее время принято делить кору на сенсорные, дви-
гательные (или моторные) и ассоциативные зоны. Такое деление было получено благодаря экспериментам на животных с экстирпацией различных участков коры, наблюдениям за больными, имеющими патологический очаг в мозге, а также с помощью прямого электрического раздражения коры
172
ипериферических структур с регистрацией электрической активности в коре.
Всенсорных зонах представлены корковые концы всех анализаторов. Для зрительного он располагается в затылочной доле мозга (поля 17, 18, 19). В поле 17 заканчивается центральный зрительный путь, информирующий о наличии и интенсивности зрительного сигнала. Это первичная зрительная область, в которой имеется топическое представительство рецепторов сетчатки, при этом каждому участку сетчатки соответствует свой участок зрительной коры. Особенно большую зону представительства имеет желтое пятно. В связи с неполным перекрестом зрительных путей в зрительную кору каждого полушария проецируются одноименные половины сетчатки обоих глаз. Это является основой бинокулярного зрения. Поля 18 и 19 – это вторичная зрительная область, которая анализирует цвет, форму, размеры и качество предмета. Нейроны этих полей полимодальны, они отвечают не только на световые, но и на тактильные, слуховые раздражители. За счет синтеза различных видов чувствительности, представленных в этих полях, возникают более сложные зрительные образы, а раздражение этих полей приводит к возникновению зрительных галлюцинаций, движений глаз и навязчивых зрительных ощущений (ауры). При поражении поля 18 больной видит, но не узнает предмета и не различает его цвета (зрительная агнозия).
Корковый конец слухового анализатора локализуется в височной доле коры (извилина Гешля), полях 41, 42, 22
ичастично 21. Они участвуют в восприятии и анализе слуховых раздражений по их громкости и тональности, организации слухового контроля речи, за который отвечает слуховой центр речи Вернике, расположенный в задних отделах верхней височной извилины (поля 22, 42, 37 левого доминантного полушария). Этот центр отвечает за распознавание и хранение устной речи как собственной, так и чужой (речевой гнозис). Больной, имеющий повреждение поля 22, теряет способность понимать значение произносимых слов. В этих полях четко представлены определенные участки кортиева ор-
173
гана (топические проекции). В средней части верхней височной извилины (поле 22) расположен центр распознавания музыкальных звуков. В поле 39 (на границе височной, теменной и затылочной долей) локализуется центр чтения письменной речи (распознавание и хранение образов письменной речи).
Ввисочной доле располагается также корковый конец
вестибулярного анализатора (задний отдел поля 21). Кожный анализатор, а также болевая, температур-
ная и проприоцептивная чувствительность проецируются на заднюю центральную извилину (поля 1, 2, 3, 43), в верхней части которой представлены нижние конечности, в средней – туловище, в нижней – руки и голова. Задняя центральная извилина вместе с прецентральной, в которой располагаются моторные зоны, объединены в так называемую «соматосенсорную кору», поскольку кожный и двигательный анализаторы тесно связаны анатомически и образуют единую функциональную единицу, обеспечивающую двигательную активность живого организма.
Вкоре теменной доли заканчиваются пути соматиче-
ской чувствительности, относящиеся к речевой функции,
связанной с оценкой воздействия на рецепторы кожи, массы
исвойств поверхности, формы и размера предмета.
Корковый конец обонятельного и вкусового анализа-
торов расположен в гиппокампальной извилине (поля 34, 43). При раздражении этой области возникают обонятельные галлюцинации, а ее повреждение приводит к аносмии (потере способности ощущать запахи).
Моторные зоны находятся в лобных долях в области передней центральной извилины мозга, раздражение которой вызывает двигательную реакцию. Кора прецентральной из-
вилины (поле 4) представляет первичную двигательную зону.
В пятом слое этого поля находятся очень крупные пирамидные клетки (гигантские клетки Беца), формирующие эфферентные выходы моторной коры на стволовые и спинальные центры. Аксоны быстропроводящих клеток Беца иннервируют крупные альфа-мотонейроны этих центров. В этой зоне
174
имеется четкая топографическая проекция мышц тела, при этом лицо проецируется на нижнюю треть прецентральной извилины, рука занимает ее среднюю треть, туловище и таз – верхнюю треть извилины. Двигательная зона коры для нижних конечностей находится на медиальной поверхности полушария в области передней части парацентральной дольки.
При поражении первичной двигательной зоны утрачивается способность к тонким координированным движениям конечностей и особенно пальцев рук.
Премоторная область коры (поле 6) располагается кпереди от первичной двигательной зоны. Поле 6 называют
вторичной моторной областью, функции которой связаны с планированием и координацией произвольных движений. Ее раздражение вызывает вращение туловища и глаз с подниманием контрлатеральной руки. Аналогичные движения наблюдаются у больных во время приступа эпилепсии, если эпилептический очаг локализуется в этой области. Поражение поля 6 у человека вызывает резкое ограничение двигательной активности, с трудом выполняются сложные комплексы движений, страдает спонтанная речь. В премоторной области коры располагается центр письменной речи (задний отдел средней лобной извилины – поле 6), центр моторной речи Брока (задний отдел нижней лобной извилины левого полушария – поле 41), а также музыкальный моторный центр (поле 45), отвечающий за тональность речи и способность петь.
К полю 6 примыкает поле 8 (лобное глазодвигательное), раздражение которого сопровождается поворотом головы и глаз в сторону, противоположную раздражаемой. Стимуляция различных участков двигательной коры вызывает сокращение соответствующих мышц на противоположной стороне.
Передние отделы лобной коры связывают с «творче-
ским» мышлением. С клинической и функциональной точек зрения интересной областью является нижняя лобная извилина (поле 44). В левом полушарии она связана с организацией двигательных механизмов речи. Раздражение этой об-
175
ласти может вызвать вокализацию, но не членораздельную речь, а также прекращение речи, если человек говорил. Поражение этой области приводит к моторной афазии – больной понимает речь, но сам говорить не может.
К ассоциативной коре относят теменно-височно- затылочную, префронтальную и лимбическую области. Она занимает около 80% всей поверхности коры больших полушарий. Ее нейроны обладают мультисенсорными функциями. В ассоциативной коре происходит интеграция различной сенсорной информации и формируется программа целенаправленного поведения, ассоциативная кора окружает каждую проекционную зону, обеспечивая взаимосвязь, например, между сенсорными и моторными областями коры. Нейроны, расположенные в этих областях, обладают полимодальностью, т.е. способностью отвечать как на сенсорную, так и моторную информацию.
Теменная ассоциативная область коры больших полу-
шарий (поля 5, 7, 40) участвует в формировании субъективного представления об окружающем пространстве, о нашем теле («схеме тела»), узнавании (гнозисе) предметов, их распознавании.
Височная область коры участвует в речевой функции посредством слухового контроля речи (слуховой центр речи Вернике). При поражении слухового центра речи больной может говорить, правильно излагать свои мысли, но не понимает чужой речи (сенсорная слуховая афазия). Эта область коры играет определенную роль в оценке пространства. Поражение зрительного центра речи приводит к потере способности читать и писать. С височной корой связывают функцию памяти и сновидений.
Лобные ассоциативные поля (поля 9–14) имеют прямое отношение к лимбическим отделам мозга, они принимают участие в формировании программы целенаправленных, сложных поведенческих актов в ответ на воздействие внешней среды на основе сенсорных сигналов всех модальностей, особенно в новой для человека обстановке. В этот процесс входят: формирование доминирующих мотиваций, обеспече-
176
ние вероятностного прогнозирования и самоконтроля действия. После префронтальной лоботомии у больных развивается так называемая «эмоциональная тупость»: отсутствие мотиваций, планирования своих действий. Этих людей отличают грубость, нетактичность, желание многократно повторять одни и те же двигательные акты.
Особенностью ассоциативной коры является пластичность нейронов, способных к перестройкам в зависимости от поступающей информации. После операции удаления какойлибо области коры в раннем детстве утраченные функции этой области полностью восстанавливаются.
Кора больших полушарий способна, в отличие от нижележащих структур мозга, длительно, в течение всей жизни, сохранять следы поступившей информации, т.е. участвовать в механизмах долговременной памяти.
Кора больших полушарий – регулятор вегетативных функций организма («кортиколизация функций»). В ней представлены все безусловные рефлексы, а также внутренние органы. Без коры невозможно выработать условные рефлексы на внутренние органы. При раздражении интерорецепторов методом вызванных потенциалов, электростимуляции и разрушения определенных участков коры доказано ее влияние на деятельность различных органов. Так, разрушение поясной извилины изменяет акт дыхания, функции сер- дечно-сосудистой системы, желудочно-кишечного тракта. Кора тормозит эмоции – «умейте властвовать собой».
Электрическая активность коры головного мозга и методы исследования функций центральной нервной системы
Мембранный потенциал пирамидных клеток составляет от 50 до 80 мкВ, потенциал действия – 60–100 мкВ. Частота ПД около 100 Гц. Он возникает в аксонном холмике нейронов коры, регистрируется с помощью микроэлектродной техники. При помещении электрода на поверхность нейрона можно записать его спонтанную, ритмическую активность.
177
С поверхности обнаженной коры между двумя электродами регистрируется суммарная электрическая активность коры и подкорковых структур, которая называется
электрокортикограммой (ЭКоГ). Эту активность можно за-
писать и с интактной кожи головы – это электроэнцефалограмма (ЭЭГ). При этом можно использовать или два активных электрода, помещенных на коже головы (биполярное отведение), или один активный, другой индифферентный (на мочке уха) (монополярное отведение).
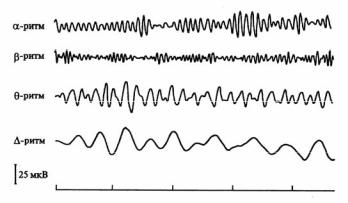
При закрытых глазах, но не во время сна регистрируется альфа-ритм, особенно четко в затылочной области, его частота 8–13 Гц, амплитуда около 50 мкВ (рис. 3.18). Действие раздражителя (например звука), переход к какой-либо деятельности при открытых глазах приводит к переходу аль- фа-ритма к бета-ритму с большей частотой (14–30 Гц) и амплитудой 25 мкВ. Это называется реакцией десинхрониза-
ции ЭЭГ.
Рис. 3.18. Ритмы электроэнцефалограммы
Переход от бодрствования ко сну сопровождается возникновением тета-ритма (частота 4–7 Гц и амплитуда 100–150 мкВ). При глубоком сне регистрируется дельтаритм с частотой 0,5–3,5 Гц. Амплитуда этих медленных волн составляет 100–300 мкВ.
178