

2 курс / Микробиология 1 кафедра / Доп. материалы / Частная бактериология
.pdf621
Рисунок 8.52 – Шарль Манту (Charles Mantoux, 1877-1947 гг.). Заимствовано из Интернет-ресурсов.
В 1912 г. австрийский ученый А. Гон (рисунок 8.53) описал специфическое туберкулезное поражение в легких – обызвествленный первичный очаг, который стали называть очагом Гона.
Рисунок 8.53 – Антон Гон (Anton Ghon, 1866-1936 гг.). Заимствовано из Интернетресурсов.
В 1919 г. французские исследователи микробиолог А. Кальметт и ветеринарный врач К. Герен (рисунок 8.54) получили вакцинный штамм микобактерий туберкулеза (бациллы Кальметта-Герена, BCG – Bacilles CalmetteGuerin или БЦЖ). Этот штамм они селекционировали путем 230 последовательных пересевов возбудителя туберкулеза бычьего типа на картофельно-глицериновой среде с добавлением желчи, затратив на это 13 лет.

622
Рисунок 8.54 – Альберт Кальметт (Leon Charles Albert Calmette, 1863-1933 гг., справа) и Камиль Герен (Jean-Marie Camille Guerin, 1872-1961 гг., слева).
Заимствовано из Интернет-ресурсов.
Во Франции 18 июня 1921 г. вакцина БЦЖ была впервые введена новорожденному ребенку. Этот день считается днем рождения БЦЖ. В 1925 г. А. Кальметт передал вакцинный штамм в СССР профессору Л.А. Тарасевичу. Через 3 года изучения было установлено, что вакцина безвредна, а смертность от туберкулеза среди вакцинированных стала намного меньше, чем среди невакцинированных людей. В 1928 г. вакцину БЦЖ стали вводить новорожденным в очагах туберкулезной инфекции. В середине 1950-х годов вакцинация новорожденных против туберкулеза в нашей стране стала обязательной.
В 1943 г. американский микробиолог З.А. Ваксман (рисунок 8.55) получил стрептомицин – первый противотуберкулезный антибиотик.
Рисунок 8.55 – Зельман Абрахам Ваксман (Selman Abraham Waksman, 1888-1973
гг.). Заимствовано из Интернет-ресурсов.

623
В 1952 г. З.А. Ваксман за открытие первого противотуберкулезного антибиотика стрептомицина был удостоен Нобелевской премии по физиологии и медицине.
Морфологические и тинкториальные свойства. Микобактерии туберкулеза имеют вид прямых или слегка изогнутых палочек длиной 1-10 мкм, диаметром 0,2-0,6 мкм (рисунок 8.56).
а б Рисунок 8.56 – Микобактерии туберкулеза: а – компьютерная визуализация; б –
сканирующая электронная микроскопия. Заимствовано из Интернет-ресурсов.
Иногда микобактерии образуют нитевидные структуры, напоминающие мицелий грибов, что и послужило основанием для их названия (рисунок 8.57).
Рисунок 8.57 – Образование микобактериями нитевидных структур, напоминающих мицелий грибов. Заимствовано из Интернет-ресурсов.
Микобактерии неподвижны, спор не образуют, имеют микрокапсулу (рисунок 8.58).
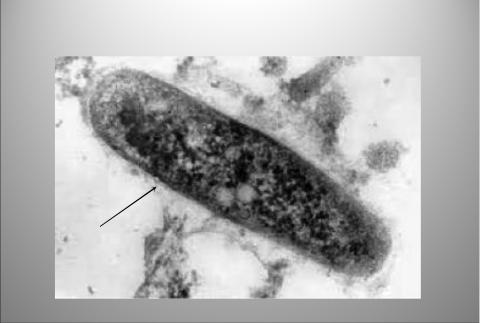
Электронная микрофотография
624
Микрокапсула
Рисунок 8.58 – Просвечивающая электронная микроскопия M. tuberculosis. Заимствовано из Интернет-ресурсов.
По химическому составу микобактерии отличаются от других микроорганизмов, они содержат специфические белки (туберкулопротеины), полисахариды и липиды. Туберкулопротеины составляют до 56% сухой массы микробной клетки. Они обладают высокой токсичностью, вызывают формирование гиперчувствительности замедленного типа (ГЗТ). Полисахариды составляют до 15% сухой массы клетки. Липиды составляют до 40% сухой массы клеток. У микобактерий обнаружено три фракции липидов: фосфатидная (растворимая в эфире), жировая (растворимая в эфире и ацетоне), восковая (растворимая в эфире и хлороформе). По структурным особенностям липиды микобактерий подразделяются на 7 групп:
-жирнокислотные производные углеводов (в частности, корд-фактор);
-маннозиды фосфатидилинозита;
-жирнокислотные производные пептидов;
-гликозиды N-ацилпептидов (микозиды С);
-жирнокислотные эфиры фтиоцеролов;
-микозиды А, В, G;
-миколаты глицерина.
Микобактерии имеют особую клеточную стенку. Наружные слои
представлены поверхностными (внешними) гликолипидами (сульфолипидами). Внешние гликолипиды называют также микозидами (специфическими восками) и сравнивают с микрокапсулой.
В средней части клеточной стенки основными компонентами являются разветвленные жирные (миколовые) кислоты с одной или двумя боковыми цепями и большим количеством атомов углерода. Миколовые кислоты обеспечивают высокую химическую устойчивость микобактерий. Миколовые кислоты могут быть ковалентно соединены с арабиногалактаном или находиться в составе свободных гликолипидов, таких как димиколат трегалозы. Наличие димиколата трегалозы способствует связыванию отдельных клеток в скопления в виде жгутов или кос, поэтому димиколат трегалозы называют “корд-фактором”.
Внутренние слои образованы арабиногалактаном и пептидогликаном. Пептидогликан непосредственно примыкает к цитоплазматической мембране.

625
Арабиногалактан связан с одной стороны с пептидогликаном, а с другой стороны – с миколовыми кислотами.
Липоарабиноманнан представляет собой смесь полимеров арабинозы и маннозы с производными пальмитиновой и туберкулостеариновой кислот. Липоарабиноманнан заякорен на цитоплазматической мембране, пронизывает клеточную стенку и выходит на ее поверхность. Концевые фрагменты липоарабиноманнана (маннозные радикалы) неспецифически подавляют активацию Т-лимфоцитов и лейкоцитов, вызывая нарушения иммунного ответа на микобактерии.
В клеточной стенке микобактерий присутствуют также маннозиды и пептиды. Поверхностные слои микобактерий из фосфолипидов, гликолипидов и миколовых кислот называют микомембраной. Клеточная стенка микобактерий пронизана порами, обеспечивающими транспорт веществ. Особое строение клеточной стенки микобактерий (как и коринебактерий) позволяет выделять кислотоустойчивый (acid fast) морфологический тип (морфотип) бактерий. Схема строения клеточной стенки микобактерий представлена на рисунке 8.59.
|
Поверхностные |
|
гликолипиды |
|
Липоарабиноманнан |
|
Миколовые кислоты |
OMP |
|
Наружная |
Наружная |
|
Арабиногалактан |
мембрана |
мембрана |
|
Пептидогликан |
MFP |
|
|
Маннозиды |
ЦПМ |
ЦПМ |
|
Цитоплазматическая |
|
мембрана |
ABC
Рисунок 8.59 – Схема строения клеточной стенки микобактерий.
Высокое содержание липидов в составе клеточной стенки определяет
спиртоустойчивость, щелочеустойчивость и кислотоустойчивость
микобактерий, затрудняет окрашивание микробных клеток обычными методами, обусловливает вирулентность и длительную сохраняемость микобактерий в окружающей среде. Кроме того, липиды экранируют бактериальную клетку, подавляют фагоцитоз, блокируют активность клеточных ферментов, а терминальные фрагменты липоарабиноманнана подавляют активацию Т- лимфоцитов и лейкоцитов.
Микобактерии относятся к группе грамположительных микроорганизмов, хотя при окраске по Граму они не прокрашиваются кристаллвиолетом. К группе грамположительных бактерий они относятся в связи с наличием слоя

626
пептидогликана, связанного с цитоплазматической мембраной и отсутствием внешней клеточной мембраны, характерной для грамотрицательных бактерий.
окраски по Цилю-
Окраска микобактерий по Цилю- |
фуксином). При |
этом |
|
Нильсену |
располагаются одиночно |
или |
|
|
рисунок 8.60). |
|
|
•Термокислотное
протравливание
карболовым фуксином. Метод предложен в 1882 г. Францем Цилем и Фридрихом Нильсеном.
Рисунок 8.60 – Окраска микобактерий по Цилю-Нельсену. Заимствовано из Интернет-ресурсов.
В цитоплазме клеток могут обнаруживаться от 2 до 12 гранул, состоящих из липидов или метафосфатов (зерна Муха).
Геном микобактерий туберкулеза представлен кольцевой молекулой ДНК (бактериальной хромосомой). Особенностью генома микобактерий туберкулеза является наличие большого количества IS-элементов, что обусловливает ДНКполиморфизм возбудителей.
Геномный анализ показал, что у микобактерий около 10% генов кодируют белки необычного аминокислотного состава. Такие белки содержат повторяющиеся сочетания либо остатков пролина и глютаминовой кислоты (РЕ-белки), либо остатков пролина, пролина и глютаминовой кислоты (РРЕ-белки). Эти белки входят в состав системы секреции VII типа (T7SS). Увеличение или снижение экспрессии генов, детерминирующих синтез этих белков, зависит от конкретных условий микроокружения. Белки семейства РЕ/РРЕ кодируются приблизительно 168 генами, что обеспечивает широкую возможность изменения их экспрессии при адаптации патогена к окружающим условиям.
Первоначально различные варианты системы секреции VII типа обозначались аббревиатурой ESX. Белки, входящие в состав этих систем, вызывают образование пор в мембранах клеток. В геноме микобактерий может присутствовать до пяти ESX-систем. Количество этих систем коррелирует с вирулентностью микобактерий. Только патогенные медленно растущие виды микобактерий имеют в своем геноме 5 ESX-систем, быстрорастущие и менее патогенные виды содержат только 3 таких кластера. Система секреции VII типа характерна для M. tuberculosis, M. bovis, M. avium. Эта система секреции состоит из двух частей (пор). Одна пора располагается в цитоплазматической мембране, а другая - в микомембране (рисунок
8.61).

627
Микомембрана
Микомембрана
Цитоплазматическая
ЦПМ
мембрана
Рисунок 8.61 – Система секреции VII типа.
Для окраски туберкулезных бактерий при люминесцентной микроскопии используют аурамин или родамин. При этом микобактерии приобретают золотистооранжевое свечение (рисунок 8.62).
Рисунок 8.62 – Люминесцентная микроскопия микобактерий. Заимствовано из Интернет-ресурсов.
Культуральные свойства. M. tuberculosis является облигатным аэробом, а M. bovis и M. africanum – аэрофилами. Оптимальная температура роста 37-38ºС. Микобактерии туберкулеза растут медленно из-за наличия в клеточной стенке липидов, замедляющих обмен веществ с окружающей средой.
Внутриклеточное дыхание микобактерий осуществляют оксидоредуктазы, из которых особый интерес представляют каталаза и пероксидаза, так как с ними связана вирулентность возбудителей.
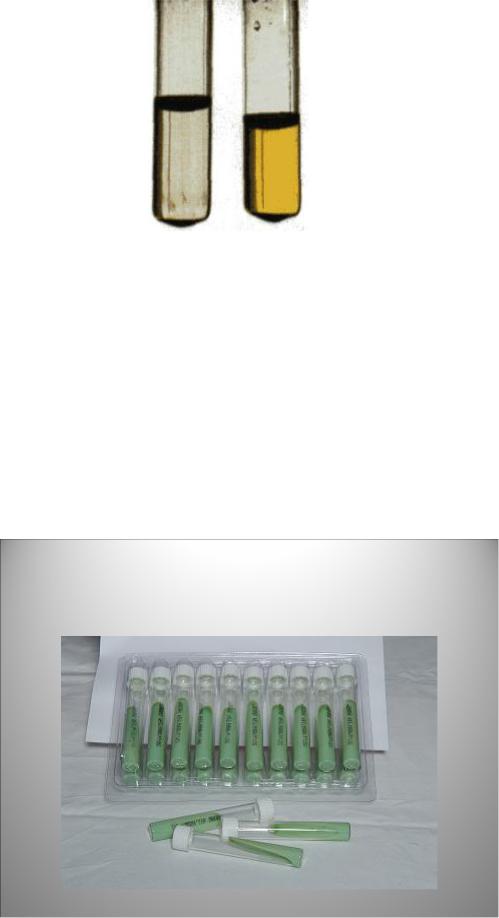
M. tuberculosis в большом количестве образует никотиновую кислоту (ниацин), которая накапливается в жидкой питательной среде и дает с раствором цианида калия и хлорамином Б ярко-желтое окрашивание - ниациновая проба Конно (рисунок 8.63).

629
Наиболее часто для выращивания микобактерий используют плотные яичные среды Левенштейна-Йенсена и Финна-2. Эти среды рекомендованы ВОЗ в качестве стандартных при диагностике туберкулеза. Они выпускаются в готовом виде в специальных пробирках с завинчивающимися пробками, предохраняющими среду от высыхания (рисунок 8.65).
Рисунок 8.65 - Коммерческая готовая среда Левенштейна-Йенсена. Заимствовано из Интернет-ресурсов.
|
На таких средах на 15-40 день культивирования микобактерии туберкулеза |
|||
образуют неправильной формы шероховатые |
Рост микобактерий на плот |
|||
(цвет “ |
|
Рост микобактерий на среде |
||
капусты |
средах в пробирках |
|||
Левенштейна-Йенсена |
||||
|
|
|
||
Рисунок 8.66 – Характер роста микобактерий туберкулеза на среде ЛевенштейнаЙенсена. Заимствовано из Интернет-ресурсов.
На средах с желчью микобактерии образуют сероватый маслянистый налет. В жидких питательных средах на 7-10 день после посева появляется пленка,
которая постепенно утолщается, становится морщинистой, приобретает желтоватый (кремовый) цвет. При этом среда остается прозрачной. При выращивании в жидкой питательной среде на стекле (метод Прайса) микобактерии туберкулеза образуют

630
структуры, напоминающие жгуты, косы, веревки. Эти структуры называются корд-фактором (рисунок 8.67).
Рисунок 8.67 – Корд-фактор M. tuberculosis - палочки расположены в виде жгутов, рисунок. Заимствовано из Интернет-ресурсов.
Резистентность микобактерий. Среди микробов, не образующих спор, микобактерии являются наиболее устойчивыми к действию неблагоприятных факторов окружающей среды. В высохшей мокроте больного микробные клетки сохраняют жизнеспособность и патогенность в течение 5-6 месяцев. При кипячении туберкулезные микобактерии погибают через 5-7 минут. На предметах больного сохраняются более 3 месяцев. В почве остаются жизнеспособными до 6 месяцев, в воде – до 15 месяцев, в навозе – 2 года. В уличной пыли микобактерии туберкулеза сохраняются в течение 10 дней. В сливочном масле микобактерии выживают до 240 дней, в сыре - до 200 дней. Солнечный свет вызывает гибель микобактерий через 1,5 часа, УФЛ – через 2-3 минуты. При пастеризации микобактерии погибают через 30 минут.
Микобактерии устойчивы к действию низких температур. Они достаточно устойчивы к действию обычных дезинфицирующих средств: 5% раствор фенола вызывает гибель туберкулезных палочек через 6 часов. Хлорсодержащие соединения (3-5% растворы хлорамина, 10-20% растворы хлорной извести), вызывают гибель возбудителя туберкулеза в течение 3-5 часов.
Эпидемиология. Основным источником инфекции при туберкулезе является больной человек. При туберкулезе легких в 50% случаев отмечается выделение возбудителя во внешнюю среду. Особое значение имеет прямой, длительный и тесный контакт здорового человека с больным. Основным
механизмом заражения является аэрогенный, а входными воротами – органы дыхания. Пути заражения туберкулезом - воздушно-капельный и воздушнопылевой (рисунок 8.68).