

6 курс / Клинические и лабораторные анализы / Кровь инфа
.pdf30
3.Функции Т-лимфоцитов:
•Распознавание антигенных детерминант (эпитопов) - обеспечивается благодаря наличию на их плазмалемме Т-клеточных рецепторов
•Элиминация антигенов - осуществляется сенсибилизированными лимфоцитами (киллерами)
•Регуляция иммунного ответа - обеспечивается специальными субпопуляциями клеток, активирующих и угнетающих иммунные реакции
•Регуляция гемопоэза (путем выделения гемопоэтических факторов)
•Регуляция пролиферации нелимфоидных клеток, участие в поддержании структурного гомеостаза (путем секреции цитокинов)
Т-хелперы
Т-хелперы участвуют и в гуморальных, и в клеточных иммунных реакциях. Причем в ходе реакции они дифференцируются в один из двух подвидов — Т-хелперы 1 или Т-хелперы 2.
Т-хелперы 1 (ТХ1) активируют |
Т-хелперы 2 (ТХ2) активируют |
|
Т-киллеры и макрофаги, |
||
В-лимфоциты и таким |
||
запуская таким образом |
||
образом значительно |
||
клеточную иммунную |
||
усиливают гуморальную |
||
реакцию и/или процесс |
||
иммунную реакцию |
||
воспаления |
||
|
В отсутствие или при недостаточной функции Т-хелперов иммунитет (особенно гуморальный) оказывается резко сниженным.

31
Распознавание антигенов и активация Т - хелперов.
Распознавание комплекса [молекулы МНС II класса + антиген], который находится на поверхности АПК, осуществляется посредством TCR и CD4 на мембране Т-хелперов. Активация Т- хелперов ведет к пролиферации соответствующего клона Т-хелперов.
Активированные Т-хелперы экспрессируют ряд костимулирующих молекул (которые усиливают взаимодействия между клетками) и выделяют лимфокины, регулирующие деятельность макрофагов, Т- и В-клеток.
Подклассы Т-хелперов (Тх1 и Тх2) различаются
характером цитокинов (лимфокинов), секретируемых при активации, экспрессией некоторых костимулирующих адгезивных молекул, а также функциональной ролью в различных звеньях иммунитета. Поддержание баланса между подклассами Т-хелперов определяет течение и исход заболеваний. Оно осуществляется благодаря тому, что клетки каждого подкласса:
•усиливают дифференцировку "наивных” Т-клеток в данный (свой) подкласс Т-хелперов. Основными аутокринными факторами роста служат γ-интерферон (для Тх1,) и интерлейкин-4 (для Тх2)
•угнетают развитие и активность клеток другого подкласса
32
Тх1 отвечают преимущественно за реакции клеточного иммунитета и воспаления, частично - за некоторые реакции гуморального иммунитета (связанные с деятельностью фагоцитов). Они выделяют ИЛ-2, ИФНγ, ФНО и экспрессируют рецепторы к ИЛ-2:
|
Интерлейкин-2 усиливает пролиферацию |
|
Т-лимфоцитов (Тх и Тк), активирует NК- |
γ-интерферон стимулирует |
клетки Т- и В-лимфоциты (под влиянием |
главным образом |
γ-интерферона переключаются на |
макрофаги |
продукцию комплемент-связывающих и |
|
опсонизирующих антител (класса IgG), |
|
которые усиливают реакции фагоцитоза) |
Фактор некроза опухоли и
γ-интерферон, привлекая и активируя гранулоциты, способствуют развитию
воспалительных реакций
Тх2 - стимулируют реакции гуморального иммунитета, участвуют в ряде регуляторных механизмов. Они выделяют интерлейкины-4, -5, -
6, -9, -10, -13, ГМ-КСФ (гранулоцитарно-макрофагальный колониестимулирующий фактор). За счет действия указанных лимфокинов происходит:
|
Активация В-лимфоцитов: усиливается их |
|
|
|
пролиферация и дифференцировка в |
Комбинация интерлейкина-4 |
|
Интерлейкин-5 усиливает |
плазматические клетки, |
||
и интерлейкина-10 усиливает |
|||
рост и активность |
вырабатывающие IgM, IgA, IgB (не |
||
рост и активность тучных |
|||
эозинофилов |
связывающие комплемент), а также IgЕ |
||
клеток и базофилов |
|||
|
(что стимулирует развитие аллергических |
||
|
|
||
|
реакций) |
|
Регуляторная функция Тх2 связана с тем, что ряд секретируемых ими цитокинов обладает противовоспалительной активностью. Вследствие этого Тх2 угнетают реакции острого и хронического воспаления. Во многих случаях это предотвращает нежелательное повреждение тканей, однако чрезмерная активность Тх2 связана с пониженным иммунитетом по отношению к внутриклеточным микроорганизмам.

33
Развитие Tx1 и Тх2 осуществляется из общего предшественника (Т-хелпера 0), причем на его преимущественное направление влияют
многочисленные факторы:
o концентрация и химическая природа антигена, пути его введения o костимулирующие адгезивные сигналы, полученные от АПК
o воздействие цитокинов, выделяемых рядом клеток микроокружения - макрофагами, лимфоцитами, тучными клетками, базофилами и эозинофилами
o генетическая предрасположенность индивидуума
Некоторые специфически активированные Т-хелперы сохраняются как долгоживущие Т-хелперы памяти, которые обеспечивают более быстрый ответ, если антиген снова появится позже.
Т-киллеры (цитотоксические Т-лимфоциты)
Т-киллеры – основные участники клеточных иммунных реакций, эти реакции развиваются в ответ на появление в организме измененных или чужеродных клеток:
•опухолевых
•трансплантированных из другого организма
•инфицированных внутриклеточными паразитами (например, вирусами)
Вбольшинстве этих случаев на клеточной поверхности оказываются чужеродные антигенные детерминанты (в комплексе с белками или в составе мембранных белков), которые могут специфически узнаваться Т-киллерами того или иного клона.
Как правило, кроме этого узнавания требуется также активация Т-киллеров другими клетками – обычно Т-хелперами.
Т-хелперы тоже должны опознать антигенные детерминанты, но способны к этому лишь при условии, что данные детерминанты находятся на поверхности не произвольных, а лишь определенных антигенпредставляющих клеток.
Активированные Т-киллеры размножаются в периферических лимфоидных органах и затем, встречая чужеродные клетки, разрушают их (механизм разрушения описывается ниже). Обломки клеток фагоцитируются макрофагами.
34
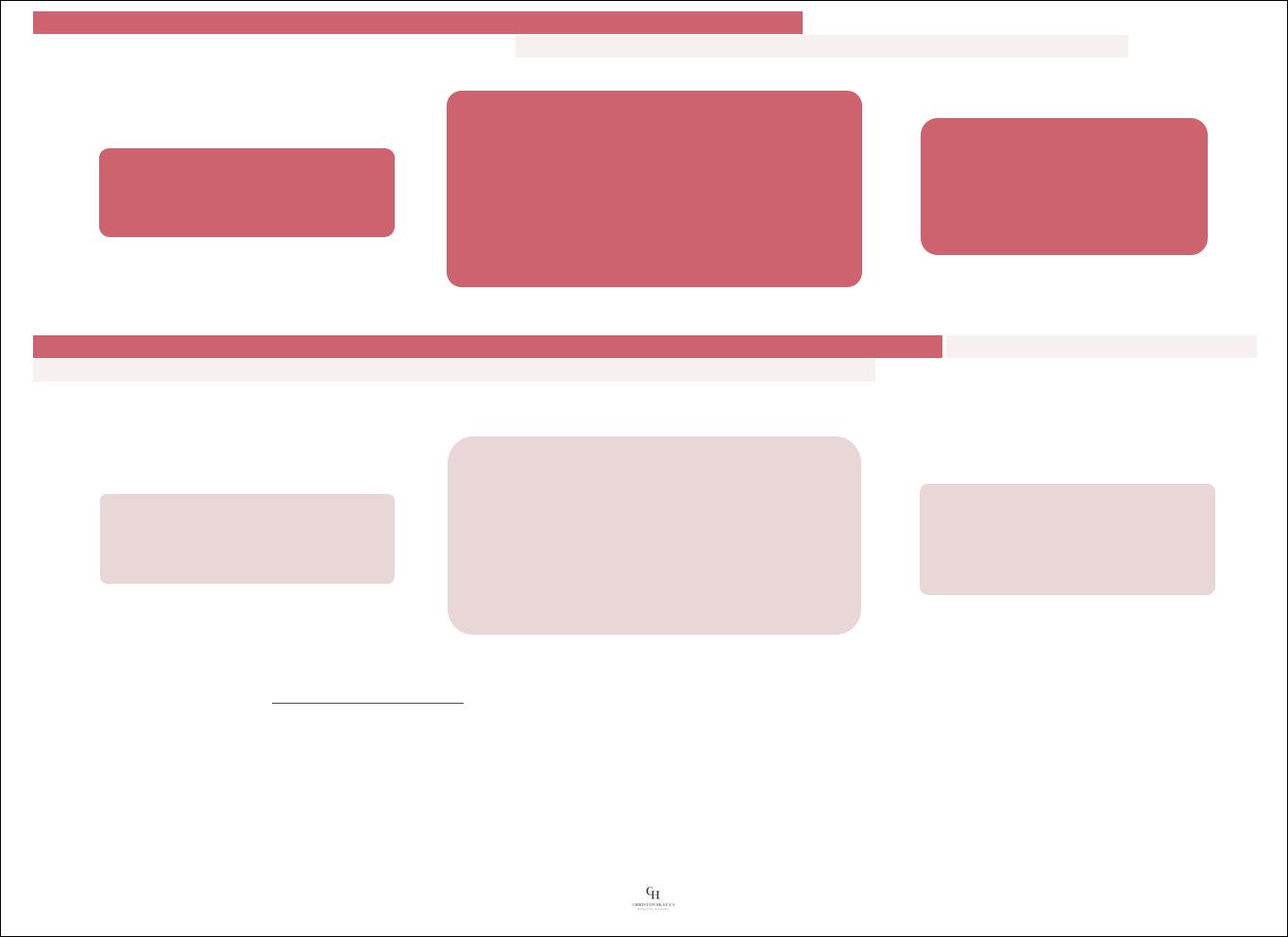
Т-киллеры посредством TCR и молекулы CD8 распознают эпитопы антигенов в комбинации с молекулами МНС I класса. Этот комплекс распознается Т-киллерами на АПК или на клеткахмишенях (измененных или чужеродных).
Распознавание антигенов и активация Т-киллеров происходит под влиянием двух основных сигналов:
1.Взаимодействия TCR и СD8 на Т-киллере с комплексом [МНС I класса + эпитоп антигена] на АПК или клетке-мишени
2.Воздействия цитокинов, выделяемых Т-хелперами, макрофагами и некоторыми АПК
Активированные Т-киллеры:
Продуцируют γ-интерферон и интерлейкин-2
Стимулируют представление антигенов, активируют клетки,
участвующие в воспалении
Уничтожают клетки-мишени, очищая от них организм
Их деятельность лежит также в основе противоопухолевого и трансплантационного иммунитета. Вещества, необходимые для уничтожения клеток-мишеней, накапливаются в образующихся 30-50 крупных цитоплазматических гранулах. По своему содержимому и морфологическим признакам эти гранулы сходны с гранулами NК-клеток.
35
При встрече с различными клетками Т-киллеры обследуют их поверхность в поисках антигенного эпитопа, который они способны распознать (в
комплексе с молекулами МНС I класса).
При обнаружении клетки-мишени зрелый Т-киллер связывается с ней и оказывает на нее летальное цитотоксическое воздействие.
После этого воздействия, осуществляемого строго прицельно и не повреждающего соседние клетки, Т-киллер отсоединяется от гибнущей
клетки-мишени и ищет следующую жертву.
 Активность Т-киллеров находится под совместным контролем Т-хелперов.
Активность Т-киллеров находится под совместным контролем Т-хелперов.
Активация Т-киллеров также приводит к образованию популяции киллерных Т-клеток памяти.
Механизмы цитотоксического действия Т-киллеров:
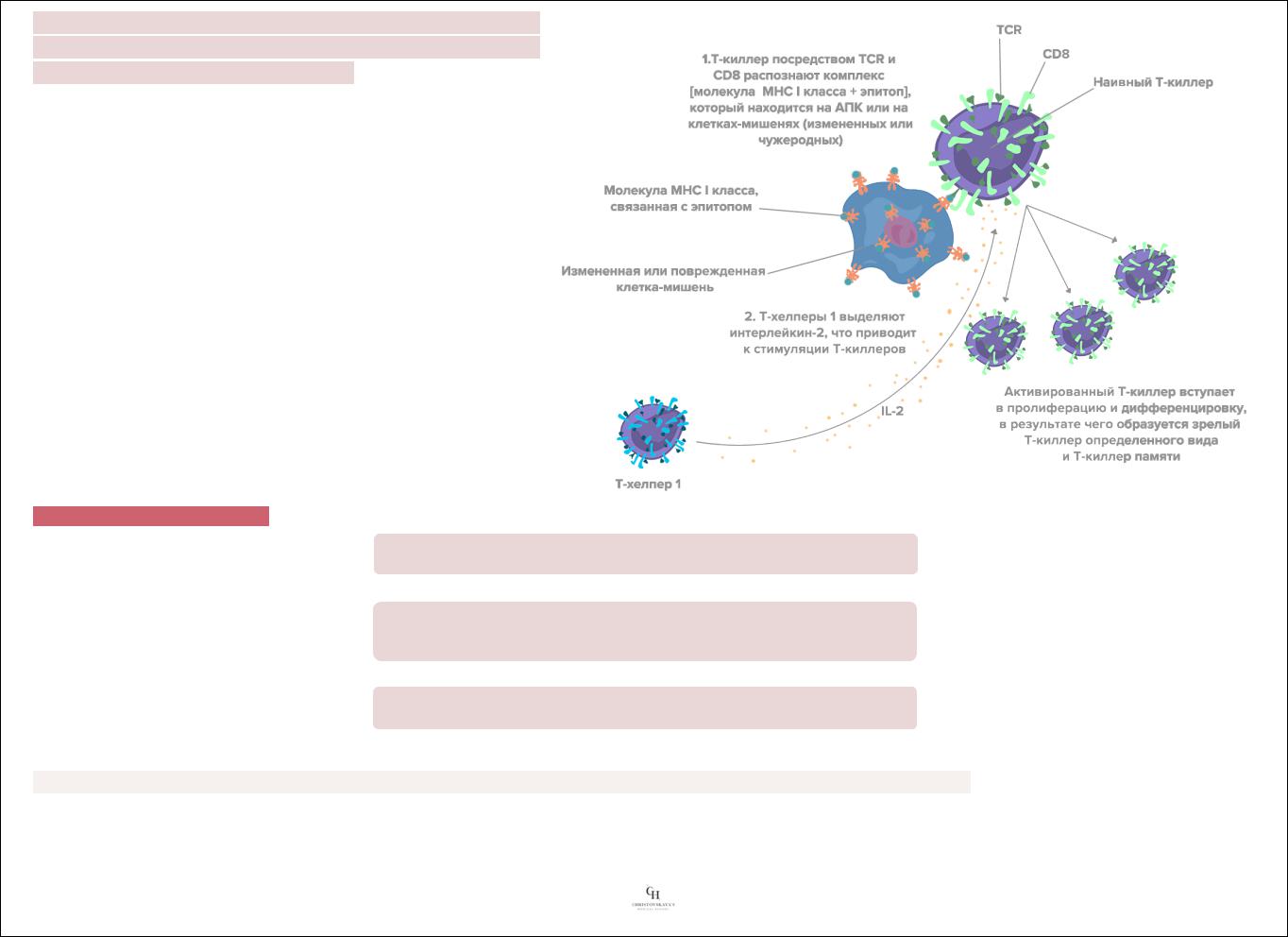
1.Образование пор в плазмолемме клеток-мишеней.
Т-киллер вступает в контакт с клеткой-мишенью и в присутствии ионов Са2+ направленно секретирует в межклеточное пространство особые белки перфорины, накопленные ранее в его цитоплазматических гранулах. Мономеры перфоринов встраиваются в качестве трансмембранных белков в плазмалемму клеток-мишеней, а в дальнейшем образуют в ней поры. Вследствие постоянного включения в мембрану новых мономеров перфоринов размеры пор растут. Формирование пор в плазмалемме приводит к осмотическому шоку клетки-мишени, ее набуханию и гибели.
36
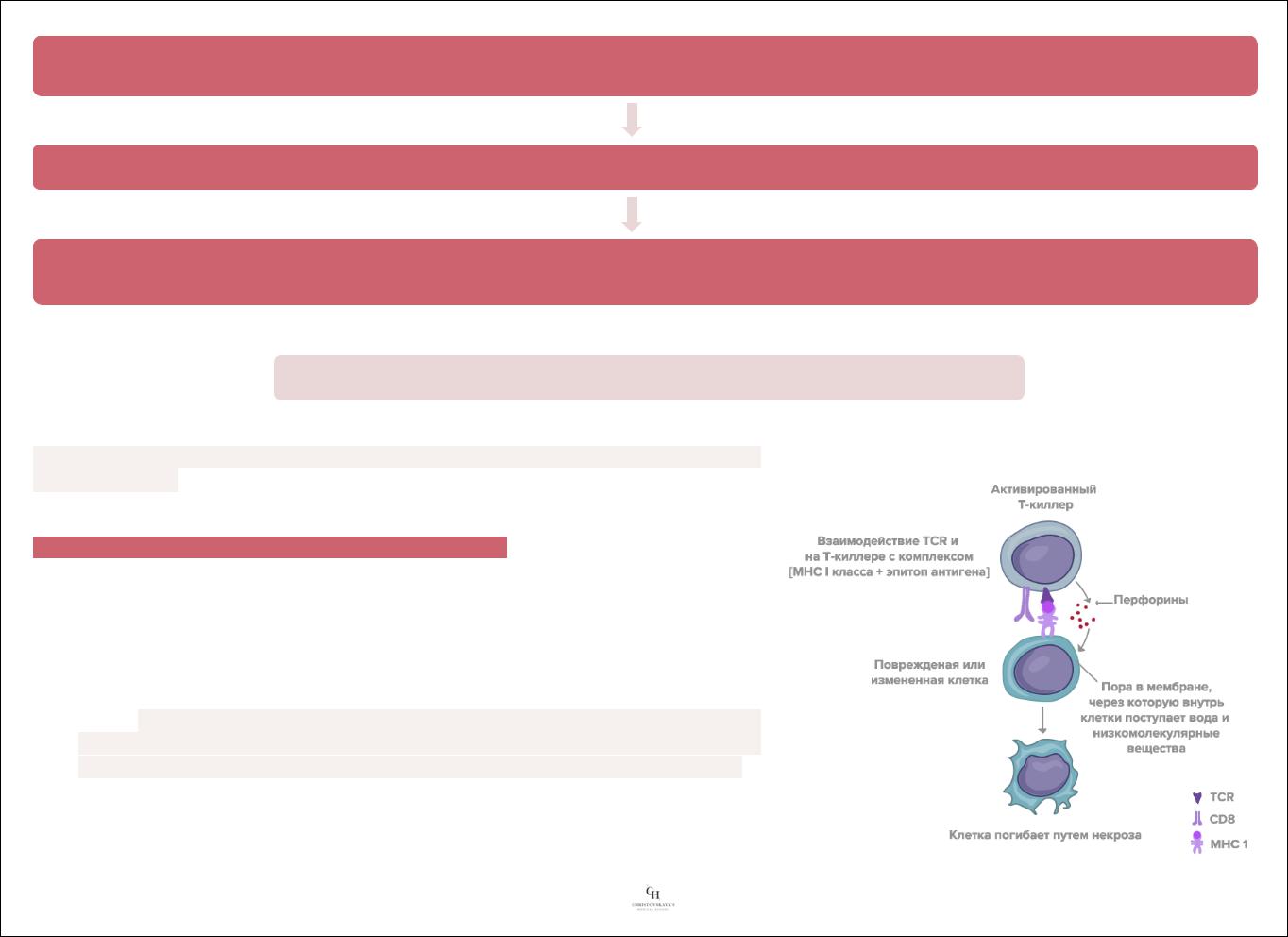
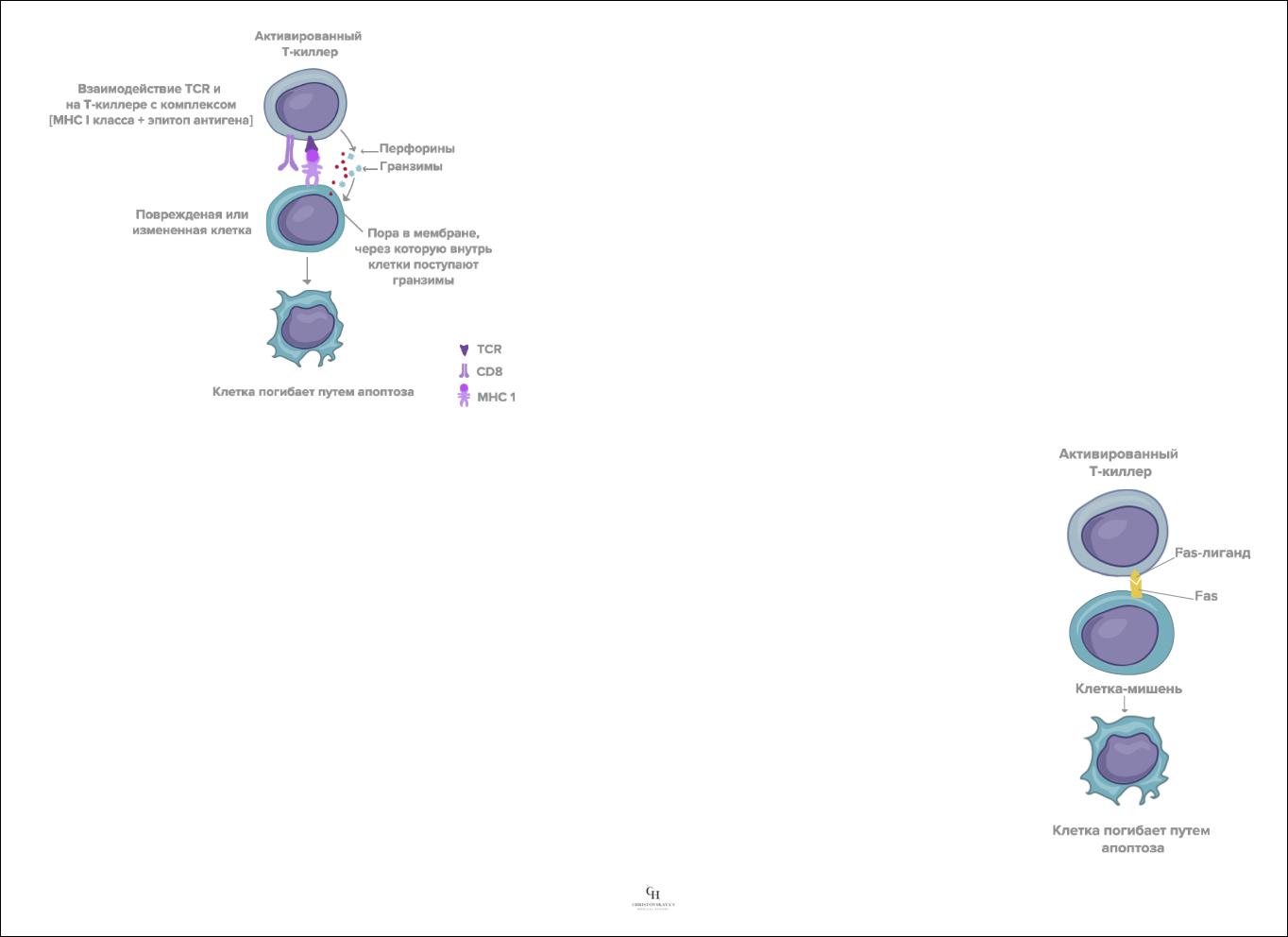
2.Индукция апоптоза клеток-мишеней ферментами, введенными в их цитоплазму через поры в плазмалемме.
Т-киллеры синтезируют и накапливают в гранулах ряд ферментов (из которых наиболее важна группа сериновых протеаз – гранзимов). Направленно выделяясь в межклеточное пространство при Са2+-зависимом экзоцитозе гранул Т-киллеров, эти ферменты через образующиеся перфориновые поры проникают в цитоплазму клеток-мишеней и запускают программу апоптоза, вызывающую гибель этих клеток.
3.Индукция апоптоза клеток-мишеней, опосредованная поверхностными рецепторами на их плазмалемме.
Происходит в результате Са2+-независимого взаимодействия антигена Fas на плазмалемме клетокмишеней с Fas-лигандом на поверхности Т-киллеров.Взаимодействие Fas и Fas-лиганда вызывает апоптоз клетки-мишени. Кроме того, это способствует также регуляции численности популяции самих Т- лимфоцитов. Активированные Т-лимфоциты резко усиливают выработку и экспрессию белка Fas (сначала неактивного) и Fas-L. По прошествии нескольких дней Fas активируется, связывается с Fas-L на этих же или других активированных лимфоцитах, вызывая запуск программы их апоптоза. Таким образом, ограничивается срок существования активированных Т-киллеров, что предотвращает возможность избыточных повреждений тканей и развития хронического воспаления.

37
Т-супрессоры
•Т-супрессоры распознают эпитопы антигенов в комбинации с молекулами МНС 1 класса.
•На их поверхности имеются TCR и молекулы CD8. Эти клетки, также идентифицируемые по наличию фактора транскрипции FOXP3, который регулирует транскрипцию генов, ответственных за дифференцировку Т-клеток и образование цитокинов, участвующих в подавлении иммунного ответа.
•Клетками-мишенями Т-супрессоров являются В-лимфоциты, Т-хелперы и Т-киллеры.
•Функция Т-супрессоров заключается в угнетении активности иммунных реакций путем непосредственного контактного воздействия на указанные клетки или секреции угнетающих (супрессорных) факторов. Т-супрессоры подавляют развитие аутоиммунных реакций.
Согласно последним исследованиям, Т-хелперы способны не только активировать, но и подавлять иммунный ответ
 (выполняют функцию супрессоров). Поэтому существование Т-супрессоров как отдельного вида Т- клеток находится под большим вопросом
(выполняют функцию супрессоров). Поэтому существование Т-супрессоров как отдельного вида Т- клеток находится под большим вопросом
Т-клетки гиперчувствительности замедленного типа (ГЗТ)
После антигенной стимуляции они начинают очень интенсивно выделять факторы, привлекающие и активирующие макрофаги, Т- киллеры и нейтрофилы, в результате чего разрушаются не только чужеродные агенты, но и клетки собственных тканей. Развивается хронический воспалительный процесс.
Т-клетки ГЗТ являются эффекторными клетками иммунных реакций гиперчувствительности замедленного типа. Они обладают поверхностным антигеном CD4 и TCR. Т-клетки ГЗТ распознают чужеродные антигены, преимущественно продуцируемые внутриклеточными микроорганизмами, в комплексе с молекулами MHC II класса. В последние годы накоплены данные, свидетельствующие о том, что роль Т-клеток ГЗТ фактически исполняют Тх1, которые и обеспечивают течение указанных реакций.
Активация Т-клеток ГЗТ сходна с активацией Т-хелперов (происходит в результате взаимодействия с АПК) и сопровождается секрецией:
•γ-интерферона, активирующего макрофаги
•МИФ (фактора, ингибирующего миграцию)
•МХФ (хемотаксического фактора макрофагов)
•других лимфокинов, которые воздействуют на макрофаги, АПК, Т-киллеры и нейтрофилы.
Таким образом, Т-клетки ГЗТ, в отличие от Т-киллеров, сами не обладают непосредственным цитотоксическим действием, однако обеспечивают защитные реакции, активно вовлекая в них клетки других типов.
Проявлением реакций ГЗТ служат очаги хронического воспаления, состоящие преимущественно из макрофагов и лимфоцитов, часто формирующих характерные компактные структуры - гранулемы. Их основу образуют скопления макрофагов (возникающие под действием МХФ и МИФ), которые находятся в активированном состоянии (эффект интерферона-γ).
38
NK-клетки: естественные киллеры или большие гранулированные лимфоциты
NK-клетки образуются в ККМ.
1)На поверхности нет Ig-подобных рецепторов. Есть рецепторы другой природы, которые настроены на узнавание определенных белков на поверхности нормальных клеток.
2)NK-клетки активируются тогда, когда указанные белки оказываются изменены (наиболее часто при опухолях и вирусах). NK-клетки – важнейший элемент противоопухолевого иммунитета.
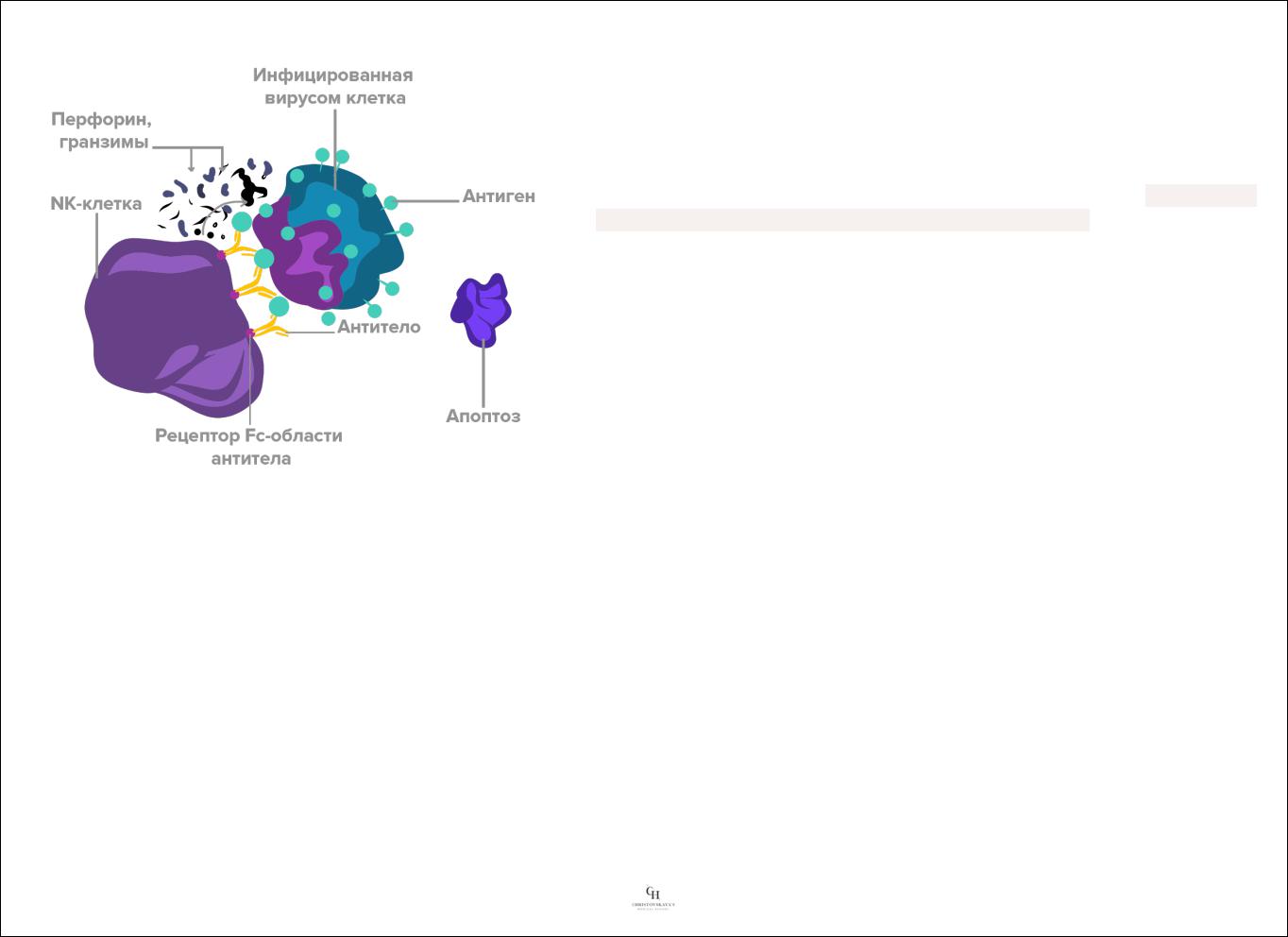
3)NК-клетки имеют рецепторы к «хвостам» IgG (Fc-фрагментам антител), благодаря чему атакуют клетки, с поверхностью которых связаны антитела.
4)В гранулах NK-клеток содержатся перфорин, протеолитические ферменты, гранзимы для разрушения клеток-мишеней (механизм такой же, как у Т-киллеров).

39
В-лимфоциты
1. Происхождение:
ККМ |
Периферические |
|
лимфоидные органы |
||
|
2. Особенности иммуноспецифичности: зрелые В-клетки имеют на поверхности
B-клеточные рецепторы (B cell receptor - BCR): В-клеточный рецептор o В-клеточные рецепторы представляют собой рецепторные
иммуноглобулины (rIg) и по антигенной специфичности последних подразделяются на множество клонов (причем у клеток любого клона содержатся rIg только одного вида)
o Специфическое взаимодействие с антигеном: когда во внеклеточной среде организма оказывается какой-либо антиген (растворенные макромолекулы или,например, бактерии),он связывается В-клетками того клона (или тех клонов), чьи BCR комплементарны данному антигену. Это первое необходимое, но не всегда достаточное условие начала гуморальной иммунной реакции.
oВ-лимфоциты экспрессируют на плазмалемме молекулы МНС I и II классов
3.Функции В-лимфоцитов:
•Распознавание антигенов иммуноглобулиновыми рецепторами
•Обеспечение реакций гуморального иммунитета
•Дифференцировка в плазмоциты - клетки, вырабатывающие иммуноглобулины